

Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase (CBS) Activity
- PMID: 35743210
- PMCID: PMC9223588
- DOI: 10.3390/ijms23126769
Identification of 8-Hydroxyquinoline Derivatives That Decrease Cystathionine Beta Synthase (CBS) Activity
Abstract
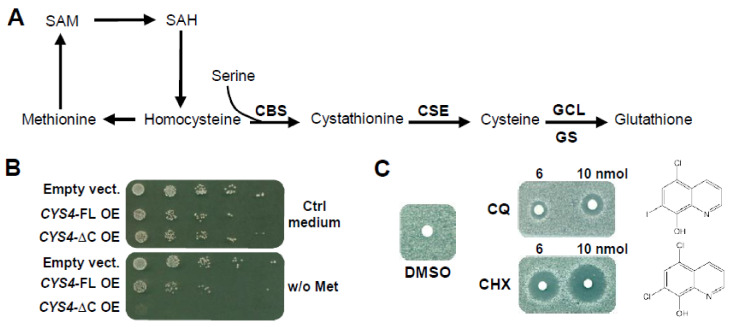
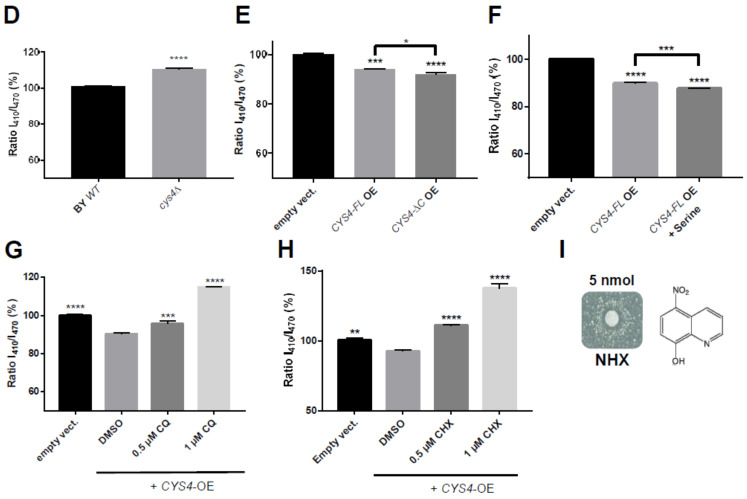
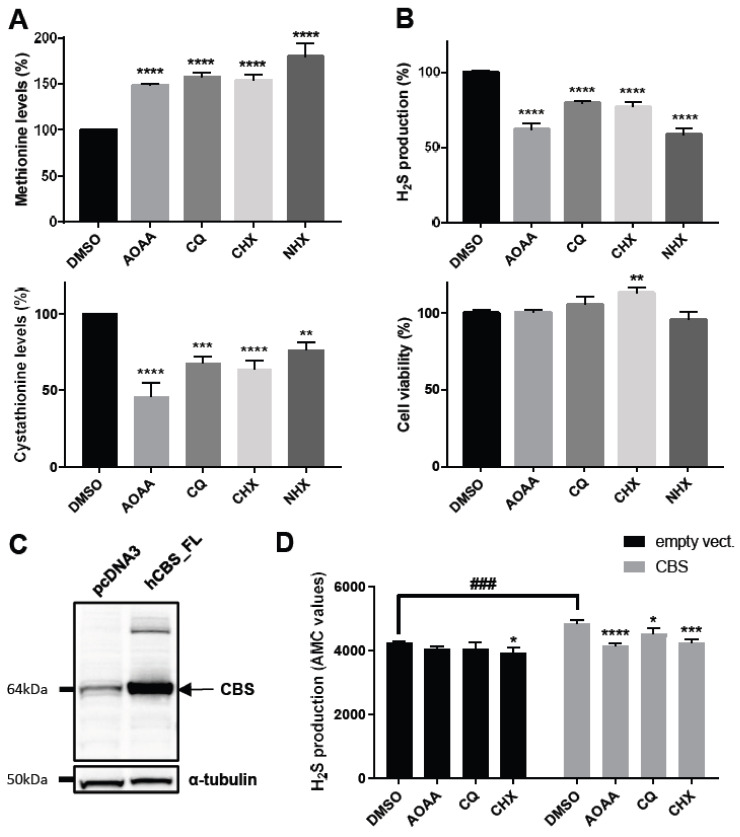
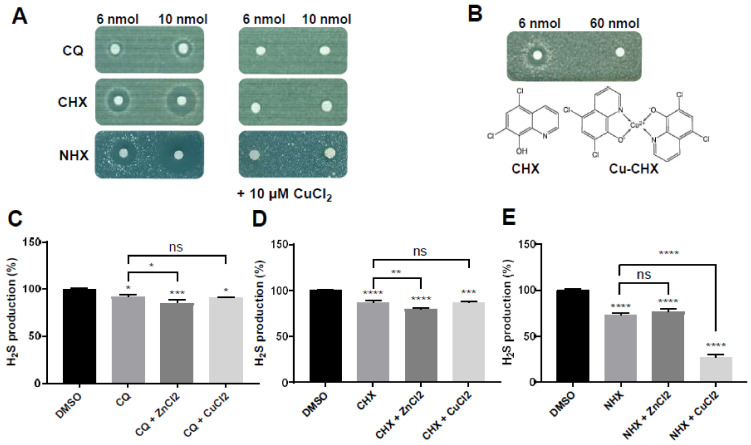
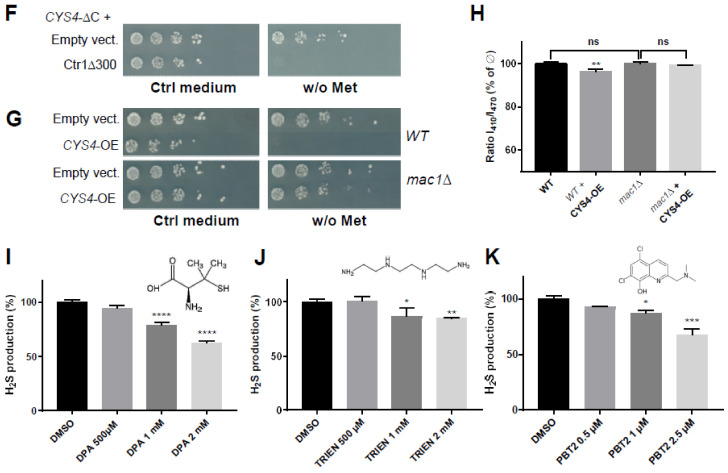
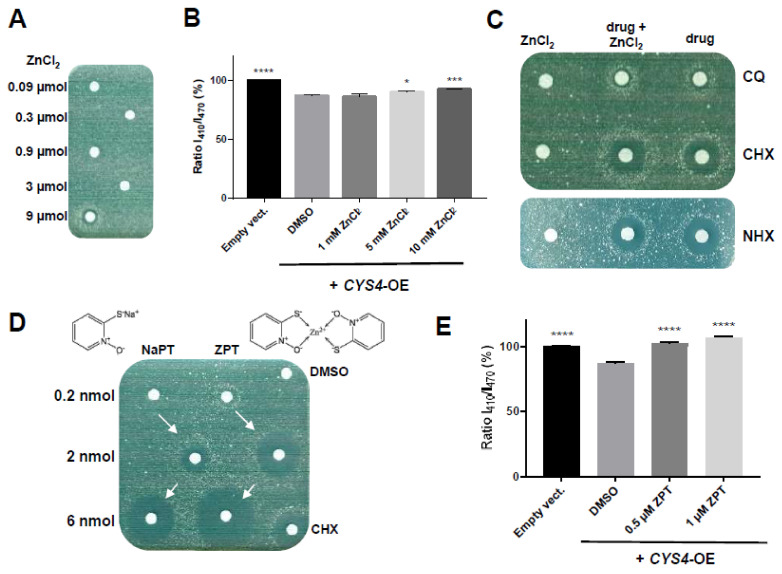
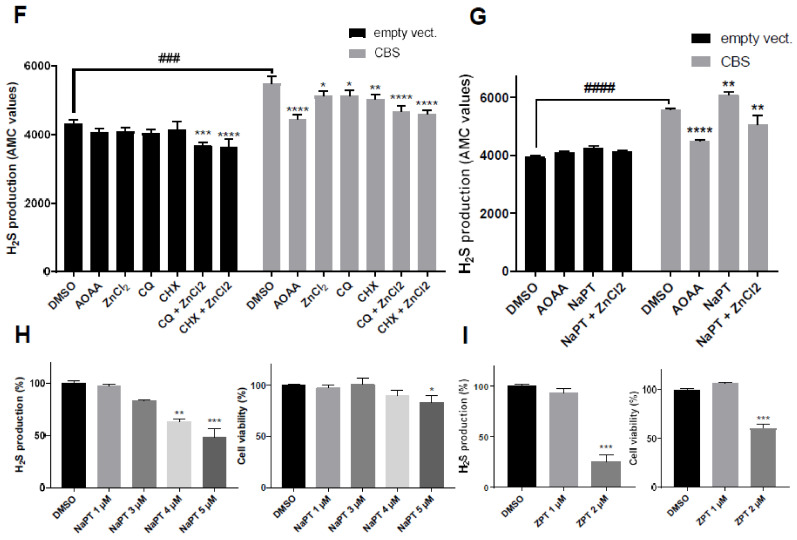
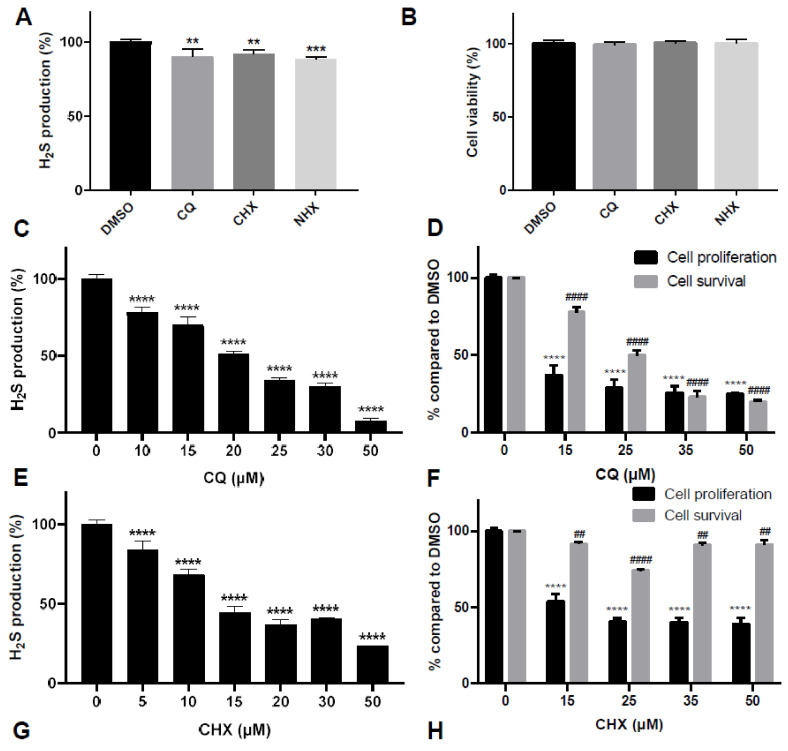
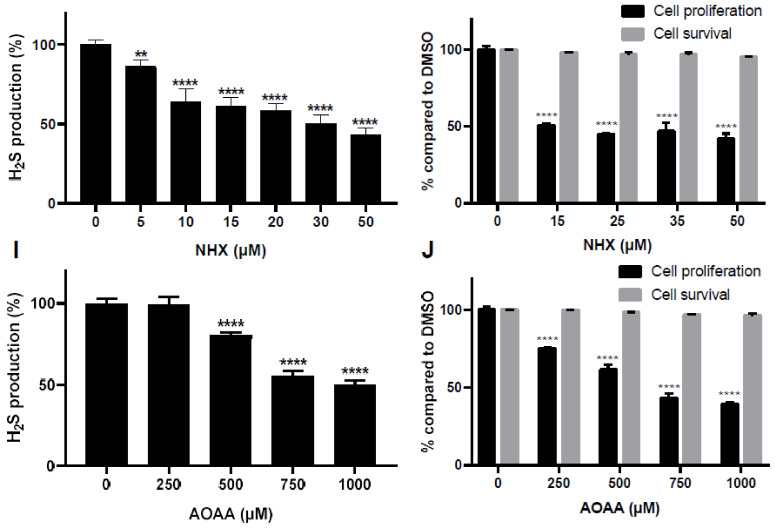
CBS encodes a pyridoxal 5'-phosphate-dependent enzyme that catalyses the condensation of homocysteine and serine to form cystathionine. Due to its implication in some cancers and in the cognitive pathophysiology of Down syndrome, the identification of pharmacological inhibitors of this enzyme is urgently required. However, thus far, attempts to identify such molecules have only led to the identification of compounds with low potency and limited selectivity. We consequently developed an original, yeast-based screening method that identified three FDA-approved drugs of the 8-hydroxyquinoline family: clioquinol, chloroxine and nitroxoline. These molecules reduce CBS enzymatic activity in different cellular models, proving that the molecular mechanisms involved in yeast phenotypic rescue are conserved in mammalian cells. A combination of genetic and chemical biology approaches also revealed the importance of copper and zinc intracellular levels in the regulation of CBS enzymatic activity-copper promoting CBS activity and zinc inhibiting its activity. Taken together, these results indicate that our effective screening approach identified three new potent CBS inhibitors and provides new findings for the regulation of CBS activity, which is crucial to develop new therapies for CBS-related human disorders.
Keywords: CBS; Cys4; Gex1/Gex2; copper; cytosolic pH; drug screening; zinc.
Conflict of interest statement
The authors have no relevant financial or non-financial interests to disclose.
Figures
References
-
- Kraus J.P., Janosík M., Kozich V., Mandell R., Shih V., Sperandeo M.P., Sebastio G., de Franchis R., Andria G., Kluijtmans L.A., et al. Cystathionine beta-synthase mutations in homocystinuria. Hum. Mutat. 1999;13:362–375. doi: 10.1002/(SICI)1098-1004(1999)13:5<362::AID-HUMU4>3.0.CO;2-K. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
