

DMT1 Protects Macrophages from Salmonella Infection by Controlling Cellular Iron Turnover and Lipocalin 2 Expression
- PMID: 35743233
- PMCID: PMC9223531
- DOI: 10.3390/ijms23126789
DMT1 Protects Macrophages from Salmonella Infection by Controlling Cellular Iron Turnover and Lipocalin 2 Expression
Abstract
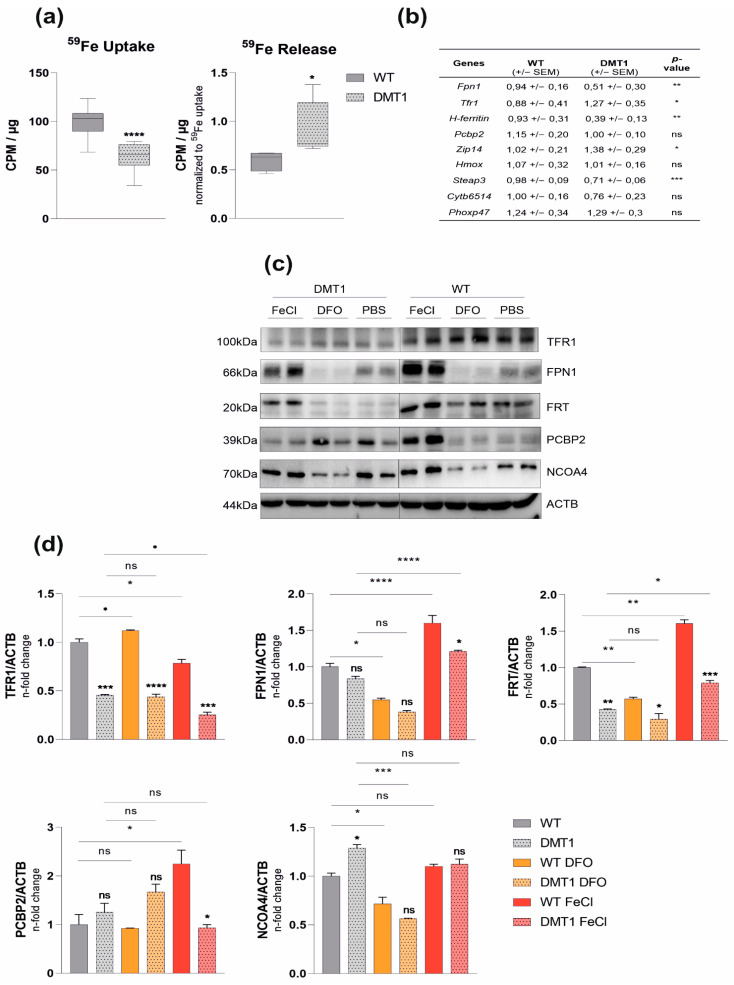
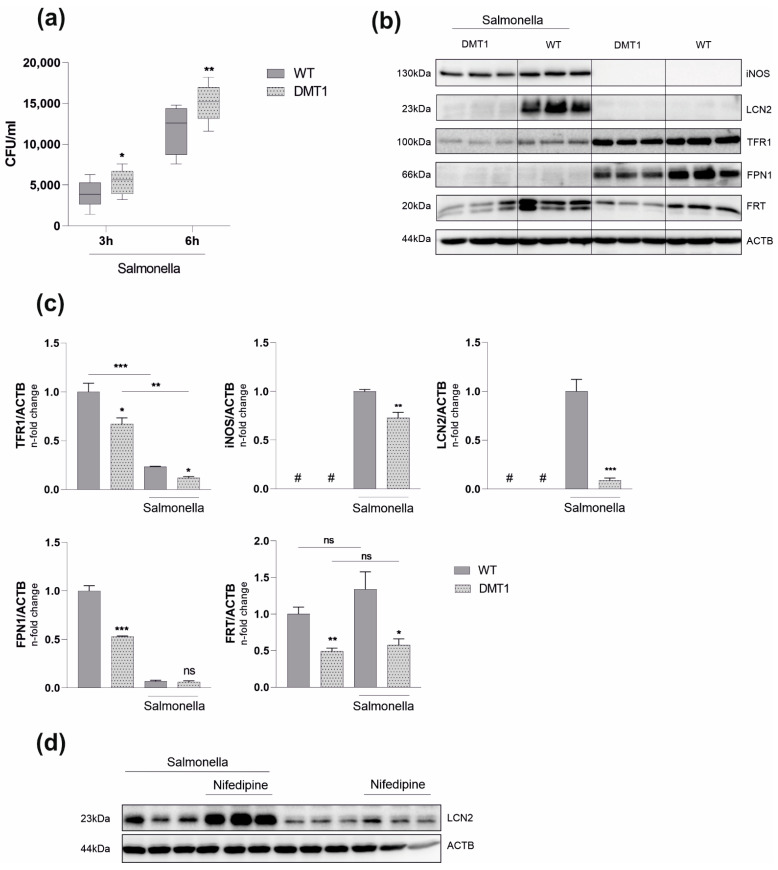
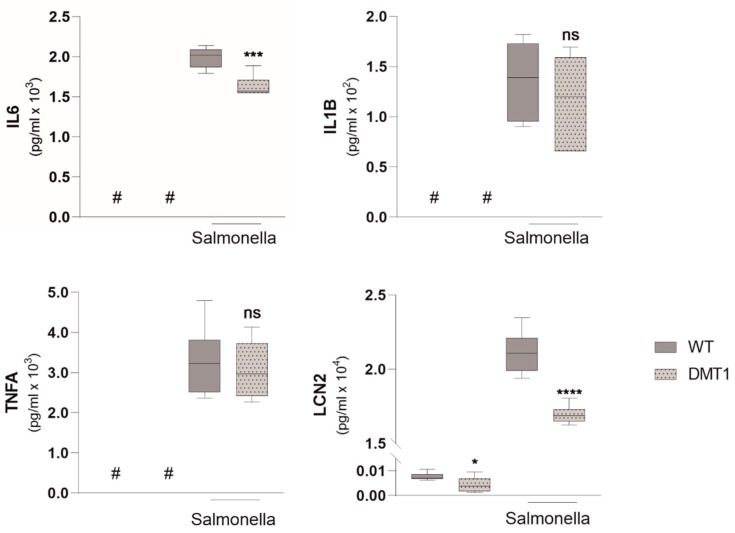
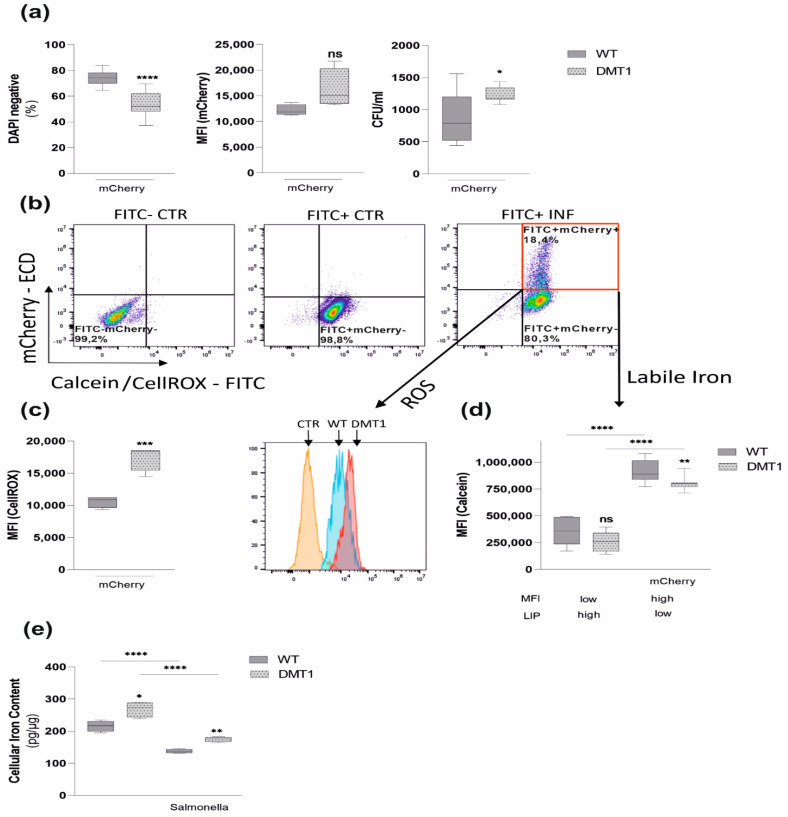
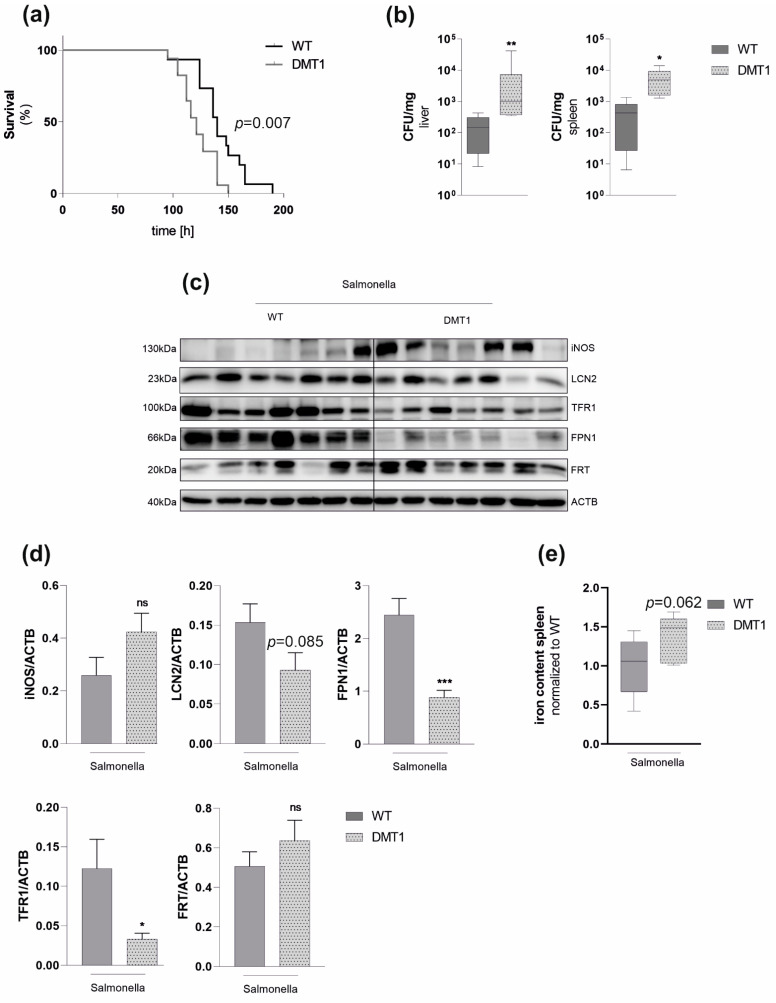
Macrophages are at the center of innate pathogen control and iron recycling. Divalent metal transporter 1 (DMT1) is essential for the uptake of non-transferrin-bound iron (NTBI) into macrophages and for the transfer of transferrin-bound iron from the endosome to the cytoplasm. As the control of cellular iron trafficking is central for the control of infection with siderophilic pathogens such as Salmonella Typhimurium, a Gram-negative bacterium residing within the phagosome of macrophages, we examined the potential role of DMT1 for infection control. Bone marrow derived macrophages lacking DMT1 (DMT1fl/flLysMCre(+)) present with reduced NTBI uptake and reduced levels of the iron storage protein ferritin, the iron exporter ferroportin and, surprisingly, of the iron uptake protein transferrin receptor. Further, DMT1-deficient macrophages have an impaired control of Salmonella Typhimurium infection, paralleled by reduced levels of the peptide lipocalin-2 (LCN2). LCN2 exerts anti-bacterial activity upon binding of microbial siderophores but also facilitates systemic and cellular hypoferremia. Remarkably, nifedipine, a pharmacological DMT1 activator, stimulates LCN2 expression in RAW264.7 macrophages, confirming its DMT1-dependent regulation. In addition, the absence of DMT1 increases the availability of iron for Salmonella upon infection and leads to increased bacterial proliferation and persistence within macrophages. Accordingly, mice harboring a macrophage-selective DMT1 disruption demonstrate reduced survival following Salmonella infection. This study highlights the importance of DMT1 in nutritional immunity and the significance of iron delivery for the control of infection with siderophilic bacteria.
Keywords: DMT1; Salmonella Typhimurium; iron; lipocalin 2; macrophage; nutritional immunity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
