

Potassium Chloroaurate-Mediated In Vitro Synthesis of Gold Nanoparticles Improved Root Growth by Crosstalk with Sucrose and Nutrient-Dependent Auxin Homeostasis in Arabidopsis thaliana
- PMID: 35745438
- PMCID: PMC9230854
- DOI: 10.3390/nano12122099
Potassium Chloroaurate-Mediated In Vitro Synthesis of Gold Nanoparticles Improved Root Growth by Crosstalk with Sucrose and Nutrient-Dependent Auxin Homeostasis in Arabidopsis thaliana
Abstract
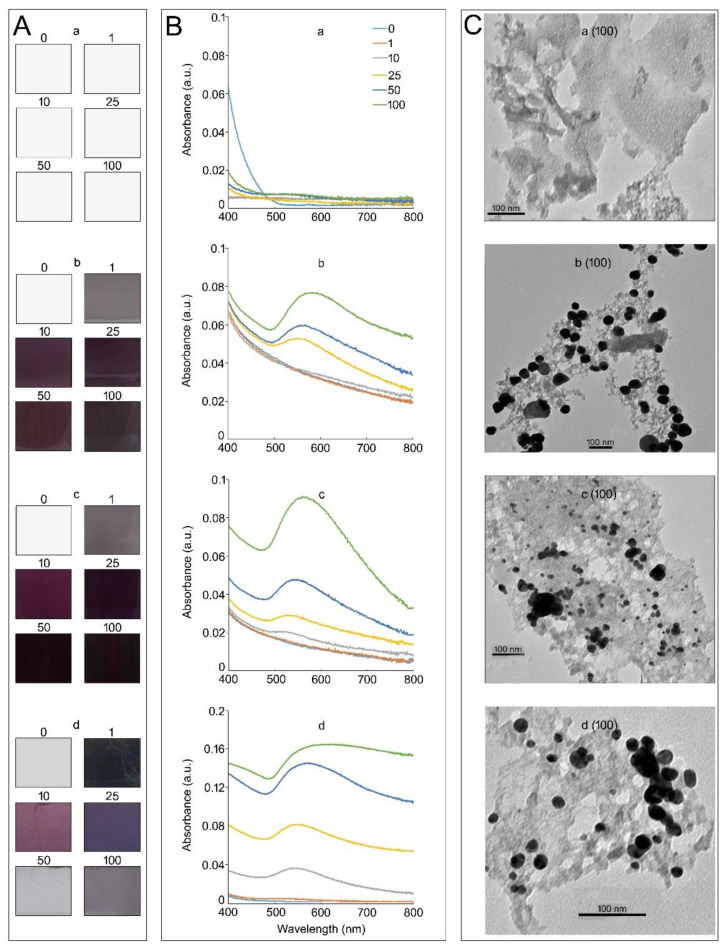
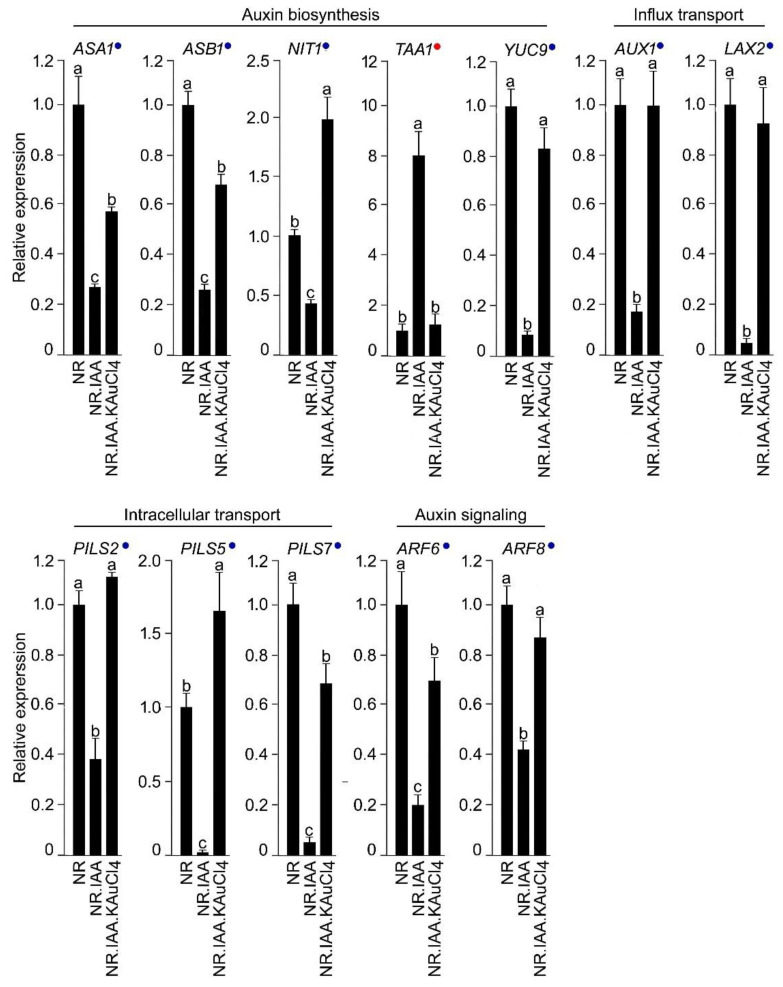
In a hydroponic system, potassium chloroaurate (KAuCl4) triggers the in vitro sucrose (Suc)-dependent formation of gold nanoparticles (AuNPs). AuNPs stimulate the growth of the root system, but their molecular mechanism has not been deciphered. The root system of Arabidopsis (Arabidopsis thaliana) exhibits developmental plasticity in response to the availability of various nutrients, Suc, and auxin. Here, we showed the roles of Suc, phosphorus (P), and nitrogen (N) in facilitating a AuNPs-mediated increase in root growth. Furthermore, the recuperating effects of KAuCl4 on the natural (IAA) auxin-mediated perturbation of the root system were demonstrated. Arabidopsis seedlings harboring the cell division marker CycB1;1::CDB-GUS provided evidence of the restoration efficacy of KAuCl4 on the IAA-mediated inhibitory effect on meristematic cell proliferation of the primary and lateral roots. Arabidopsis harboring synthetic auxin DR5rev::GFP exhibited a reinstating effect of KAuCl4 on IAA-mediated aberration in auxin subcellular localization in the root. KAuCl4 also exerted significant and differential recuperating effects on the IAA-mediated altered expression of the genes involved in auxin signaling and biosynthetic pathways in roots. Our results highlight the crosstalk between KAuCl4-mediated improved root growth and Suc and nutrient-dependent auxin homeostasis in Arabidopsis.
Keywords: Arabidopsis; KAuCl4; auxin; gold nanoparticles; hydroponic system; nutrients; sucrose.
Conflict of interest statement
The authors declare no conflict to interest.
Figures








References
-
- Bhattacharya T., Das D., Borges e Soares G.A., Chakrabarti P., Ai Z., Chopra H., Hasan M.A., Simona Cavalu S. Novel green approaches for the preparation of gold nanoparticles and their promising potential in oncology. Processes. 2022;10:426. doi: 10.3390/pr10020426. - DOI
-
- Ebrahimian J., Khayatkashani M., Soltani N., Yousif Q.A., Salavati-Niasari M. Catechin mediated green synthesis of au nanoparticles: Experimental and theoretical approaches to the determination HOMO-LUMO energy gap and reactivity indexes for the (+)-epicatechin (2S, 3S) Arab. J. Chem. 2022;15:103758. doi: 10.1016/j.arabjc.2022.103758. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
