

Genome-wide CRISPR screens of T cell exhaustion identify chromatin remodeling factors that limit T cell persistence
- PMID: 35750052
- PMCID: PMC9949532
- DOI: 10.1016/j.ccell.2022.06.001
Genome-wide CRISPR screens of T cell exhaustion identify chromatin remodeling factors that limit T cell persistence
Abstract
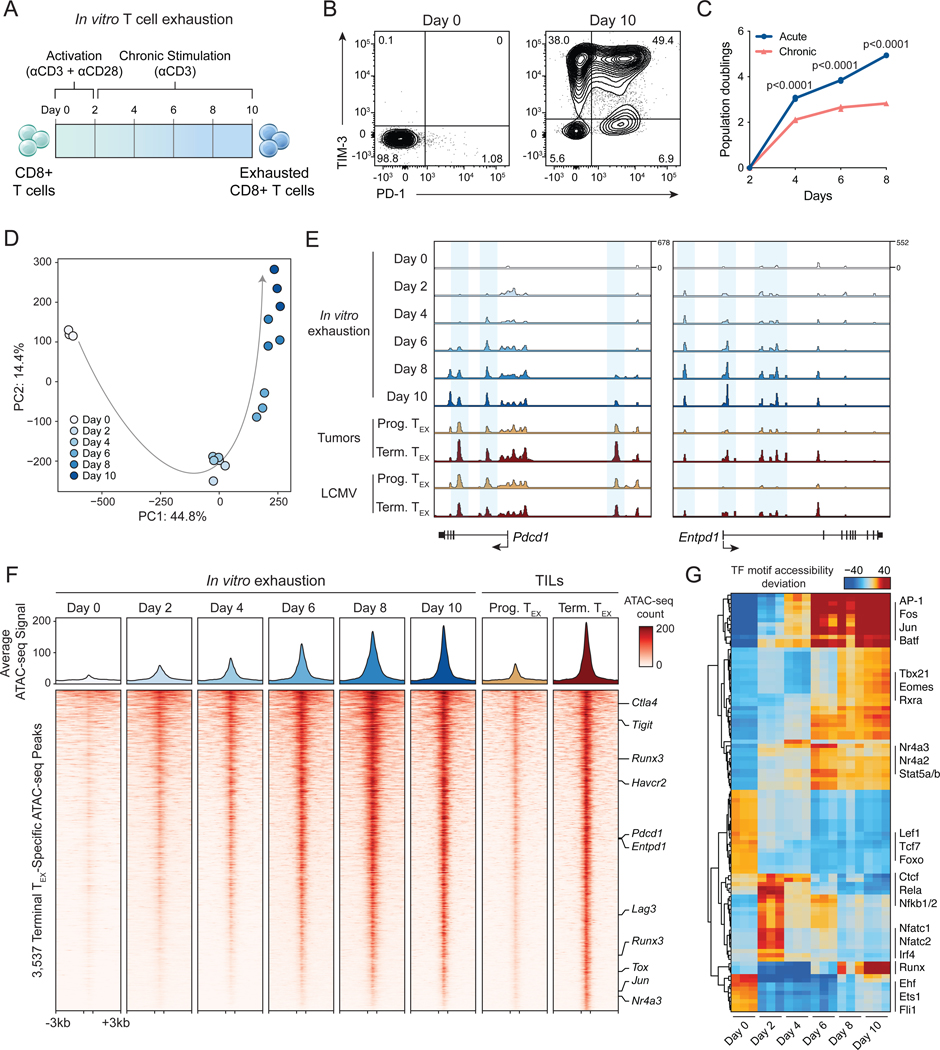
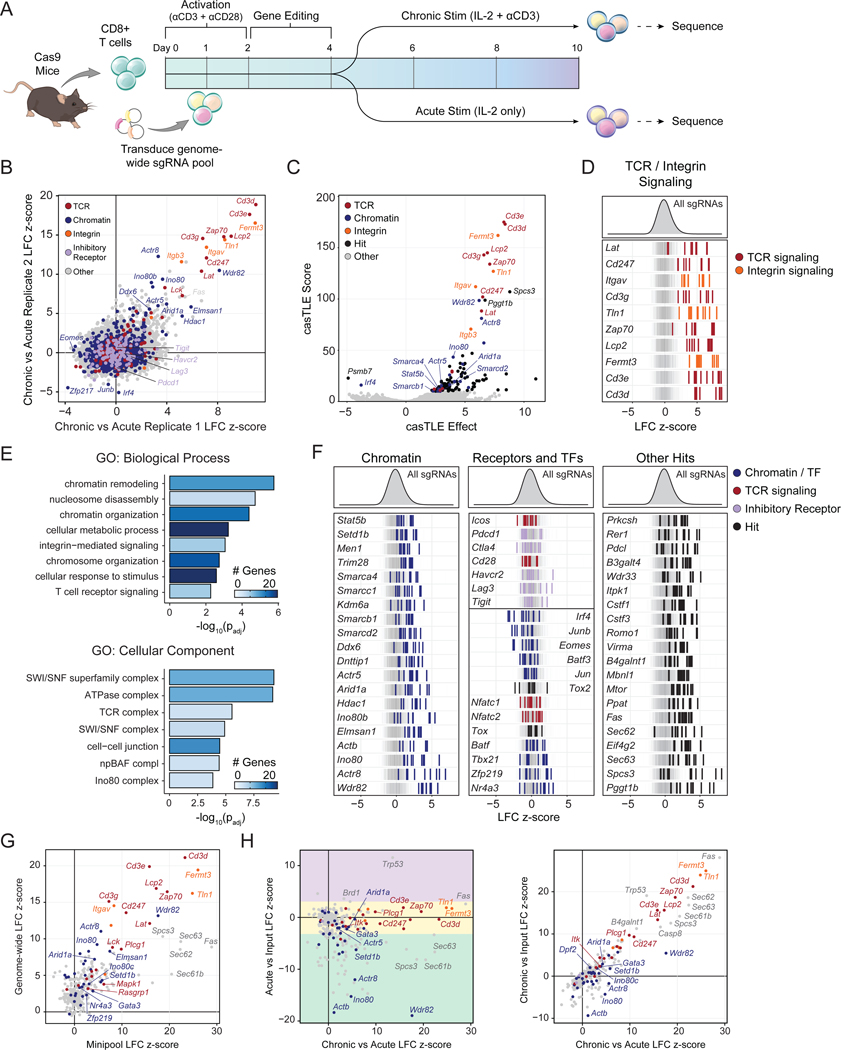
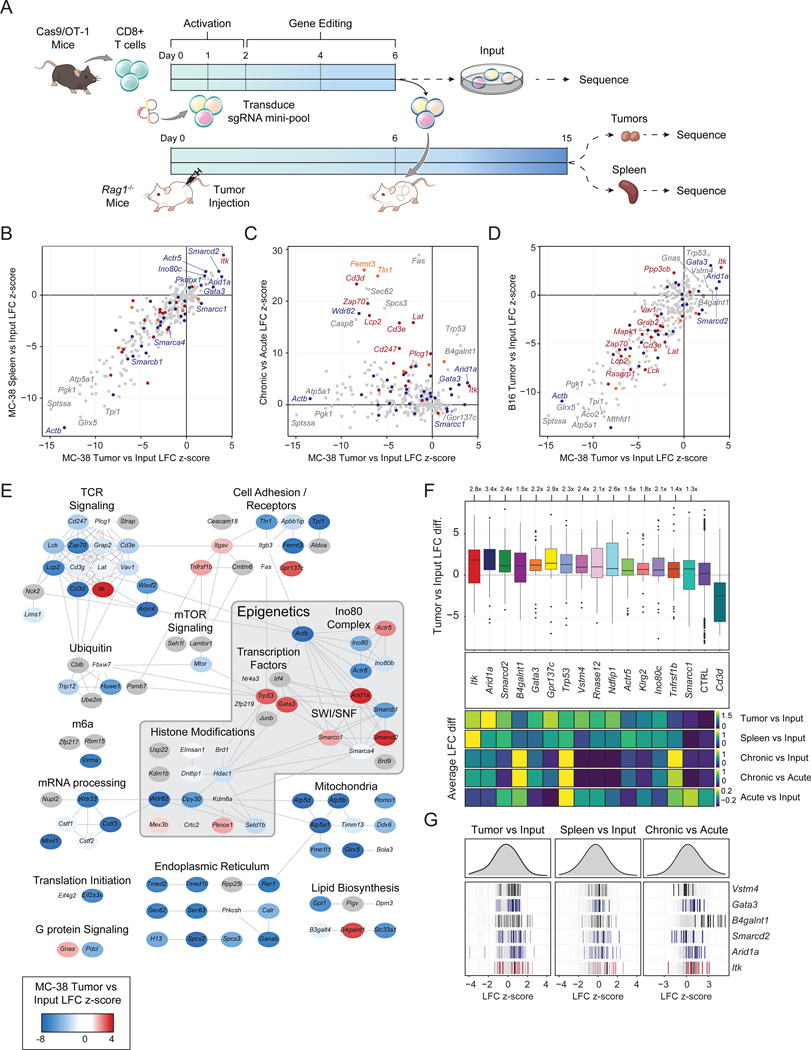
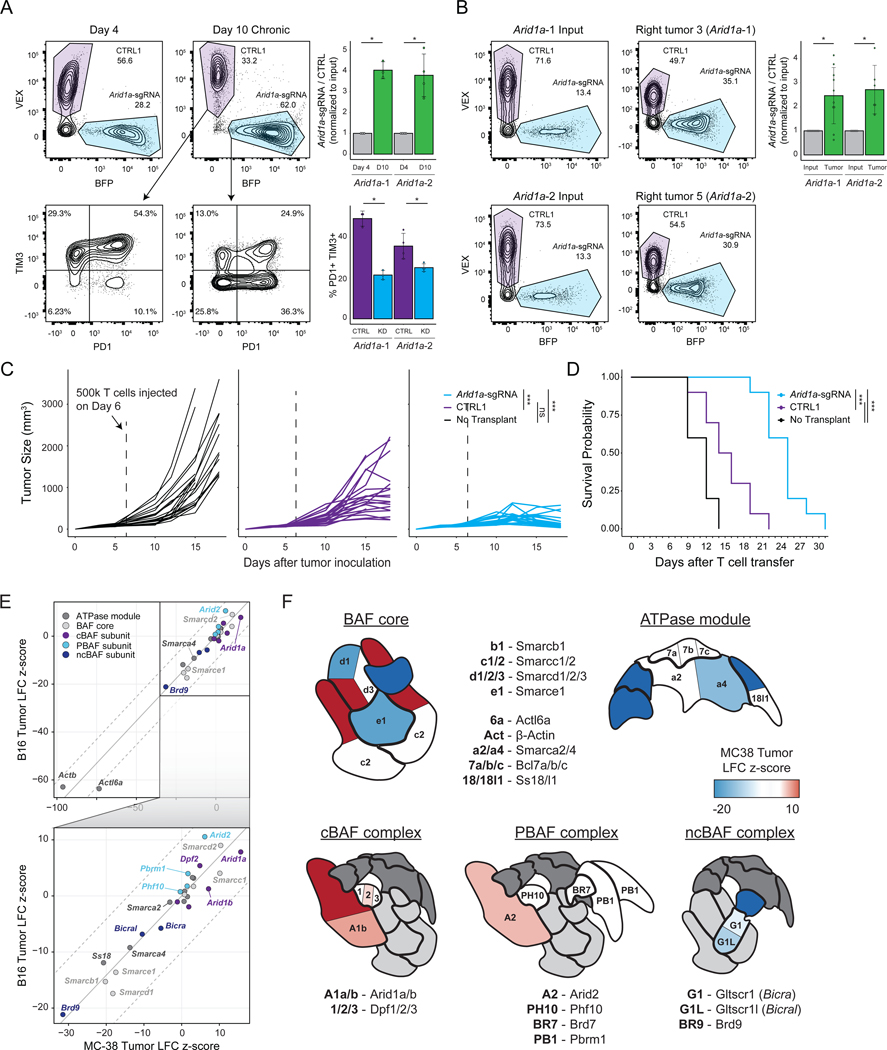
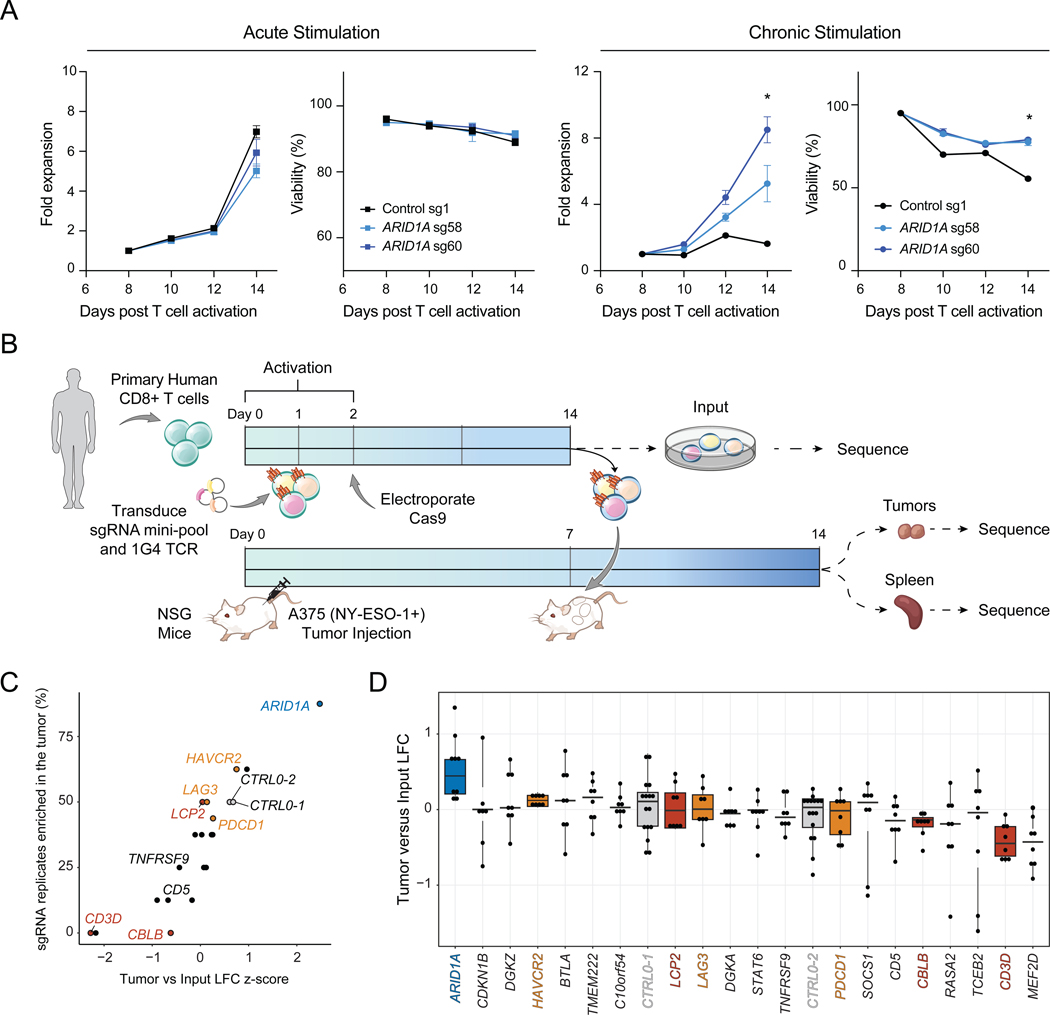
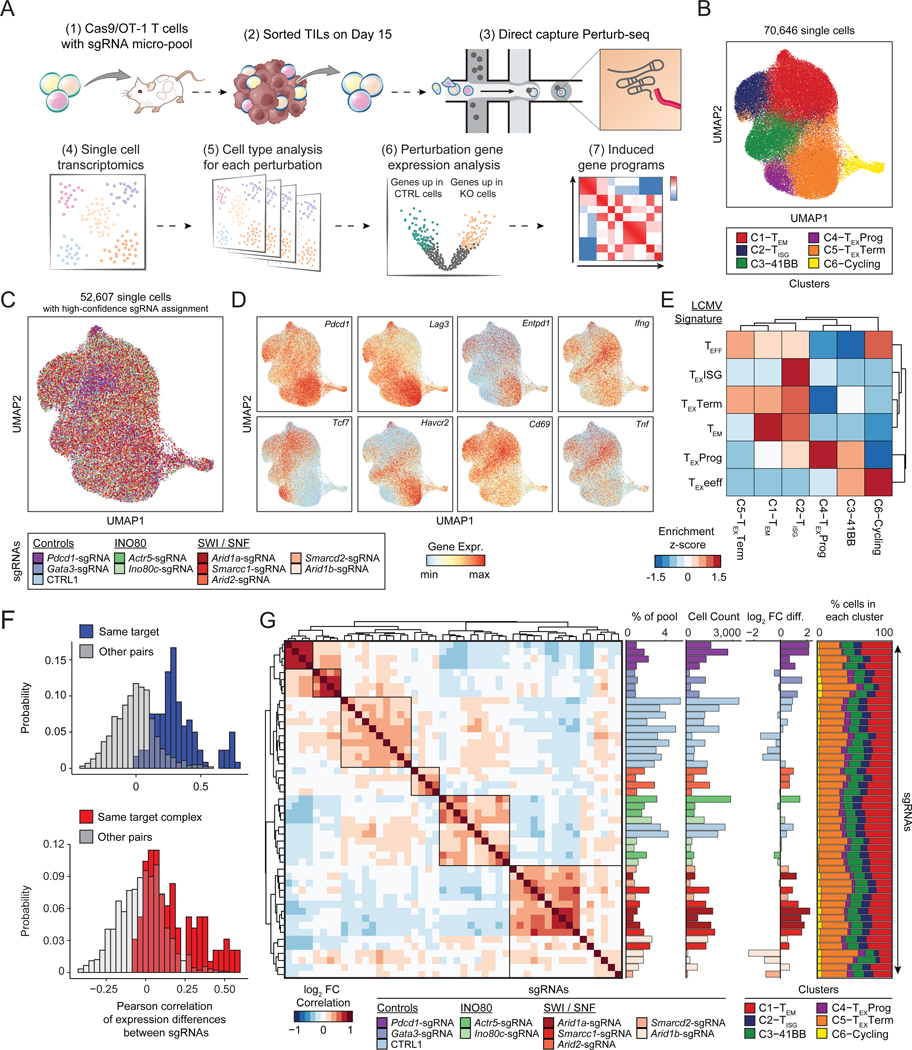
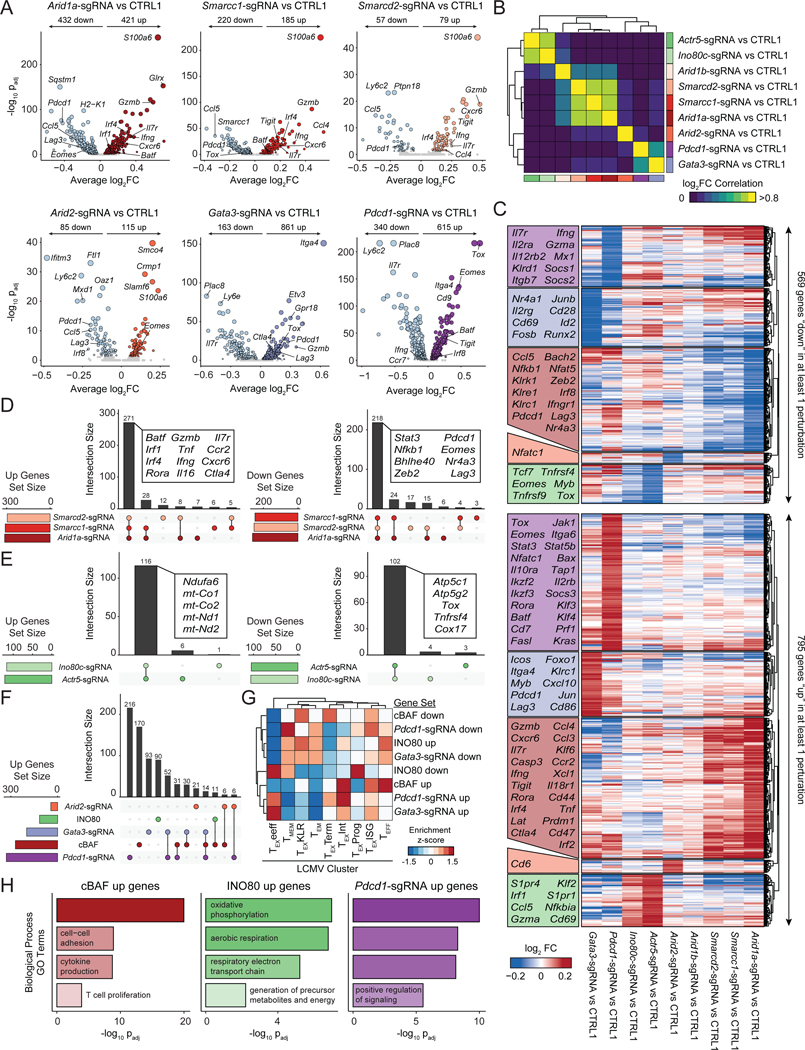
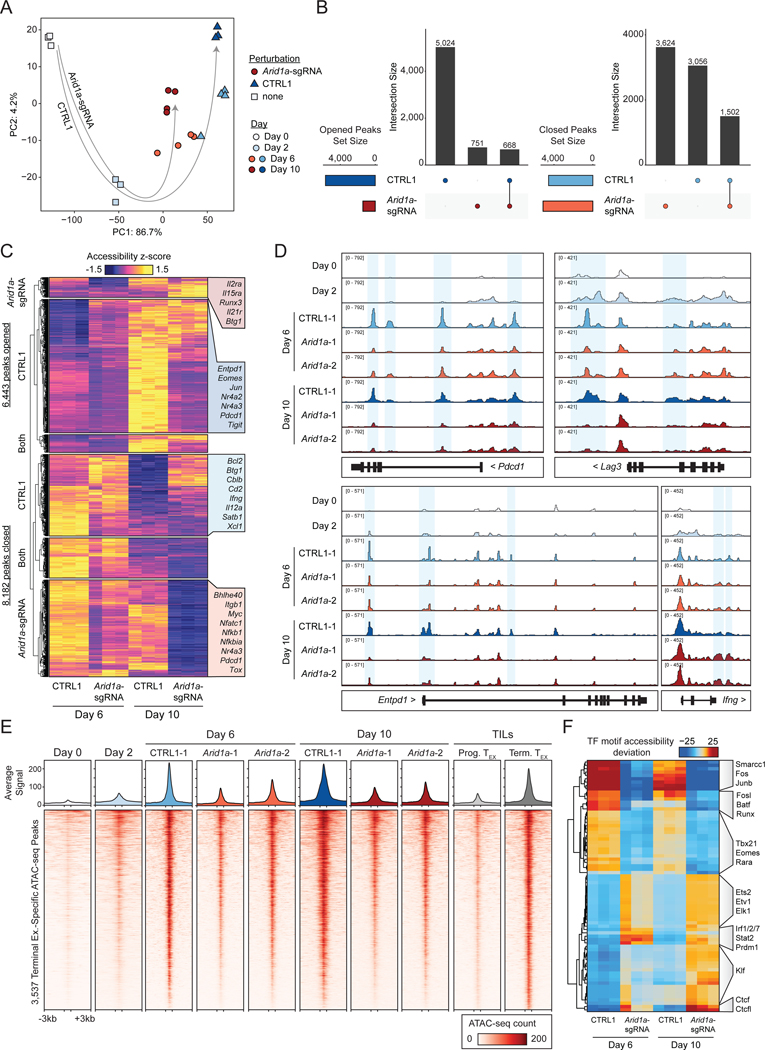
T cell exhaustion limits antitumor immunity, but the molecular determinants of this process remain poorly understood. Using a chronic stimulation assay, we performed genome-wide CRISPR-Cas9 screens to systematically discover regulators of T cell exhaustion, which identified an enrichment of epigenetic factors. In vivo CRISPR screens in murine and human tumor models demonstrated that perturbation of the INO80 and BAF chromatin remodeling complexes improved T cell persistence in tumors. In vivo Perturb-seq revealed distinct transcriptional roles of each complex and that depletion of canonical BAF complex members, including Arid1a, resulted in the maintenance of an effector program and downregulation of exhaustion-related genes in tumor-infiltrating T cells. Finally, Arid1a depletion limited the acquisition of exhaustion-associated chromatin accessibility and led to improved antitumor immunity. In summary, we provide an atlas of the genetic regulators of T cell exhaustion and demonstrate that modulation of epigenetic state can improve T cell responses in cancer immunotherapy.
Keywords: CRISPR; T cell exhaustion; canonical BAF complex; chromatin remodeling; epigenetics; genomics; immunology; in vivo Perturb-seq.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.T.S. is a scientific founder of Immunai and founder of Cartography Biosciences and receives research funding from Arsenal Biosciences, Allogene Therapeutics, and Merck Research Laboratories. J.A.B. is a consultant to Immunai. S.A.V. is an advisor to Immunai. K.E.Y. is a consultant to Cartography Biosciences. C.L.M. is a co-founder of Lyell Immunopharma and Syncopation Life Sciences, and consults for Lyell, Syncopation, NeoImmune Tech, Apricity, Nektar, Immatics, Mammoth, and Ensoma. A.A. is a co-founder of Tango Therapeutics, Azkarra Therapeutics, Ovibio Corporation, and Kytarro; a consultant for SPARC, Bluestar, Pro-Lynx, Earli, Cura, GenVivo, Ambagon, Phoenix Molecular Designs, and GlaxoSmithKline (GSK); a member of the Scientific Advisory Board of Genentech, GLAdiator, Circle and Cambridge Science Corporation; receives research support from SPARC and AstraZeneca; holds patents on the use of poly (ADP-ribose) polymerase (PARP) inhibitors held jointly with AstraZeneca. A.M. is a co-founder of Spotlight Therapeutics, Arsenal Biosciences, and Survey Genomics; a member of the Scientific Advisory Board of NewLimit; owns stock in Arsenal Biosciences, Spotlight Therapeutics, NewLimit, Survey Genomics, PACT Pharma, and Merck; has received fees from 23andMe, PACT Pharma, Juno Therapeutics, Trizell, Vertex, Merck, Amgen, Genentech, AlphaSights, Rupert Case Management, Bernstein, and ALDA; is an investor in and informal advisor to Offline Ventures; and is a client of EPIQ. The Marson lab has received research support from Juno Therapeutics, Epinomics, Sanofi, GSK, Gilead, and Anthem. K.A.F., E.S., J.C., A.A., A.M., and C.L.M. hold patents in the arena of CAR-T cell therapeutics. J.A.B. and A.T.S. have filed a patent related to the contents of this study.
Figures
Comment in
-
Chromatin Remodeling Factors Drive Epigenetic Changes Crucial for T-cell Exhaustion.Cancer Discov. 2022 Aug 5;12(8):1837. doi: 10.1158/2159-8290.CD-RW2022-121. Cancer Discov. 2022. PMID: 35775817
-
New epigenetic regulators of T cell exhaustion.Cancer Cell. 2022 Jul 11;40(7):708-710. doi: 10.1016/j.ccell.2022.06.008. Cancer Cell. 2022. PMID: 35820395
References
-
- Adamson B, Norman TM, Jost M, Cho MY, Nuñez JK, Chen Y, Villalta JE, Gilbert LA, Horlbeck MA, Hein MY, Pak RA, Gray AN, Gross CA, Dixit A, Parnas O, Regev A, Weissman JS, 2016. A Multiplexed Single-Cell CRISPR Screening Platform Enables Systematic Dissection of the Unfolded Protein Response. Cell 167, 1867–1882.e21. 10.1016/j.cell.2016.11.048 - DOI - PMC - PubMed
-
- Alfei F, Kanev K, Hofmann M, Wu M, Ghoneim HE, Roelli P, Utzschneider DT, von Hoesslin M, Cullen JG, Fan Y, Eisenberg V, Wohlleber D, Steiger K, Merkler D, Delorenzi M, Knolle PA, Cohen CJ, Thimme R, Youngblood B, Zehn D, 2019. TOX reinforces the phenotype and longevity of exhausted T cells in chronic viral infection. Nature 571, 265–269. 10.1038/s41586-019-1326-9 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
