

Seasonal influenza vaccination expands hemagglutinin-specific antibody breadth to older and future A/H3N2 viruses
- PMID: 35750781
- PMCID: PMC9232600
- DOI: 10.1038/s41541-022-00490-0
Seasonal influenza vaccination expands hemagglutinin-specific antibody breadth to older and future A/H3N2 viruses
Abstract
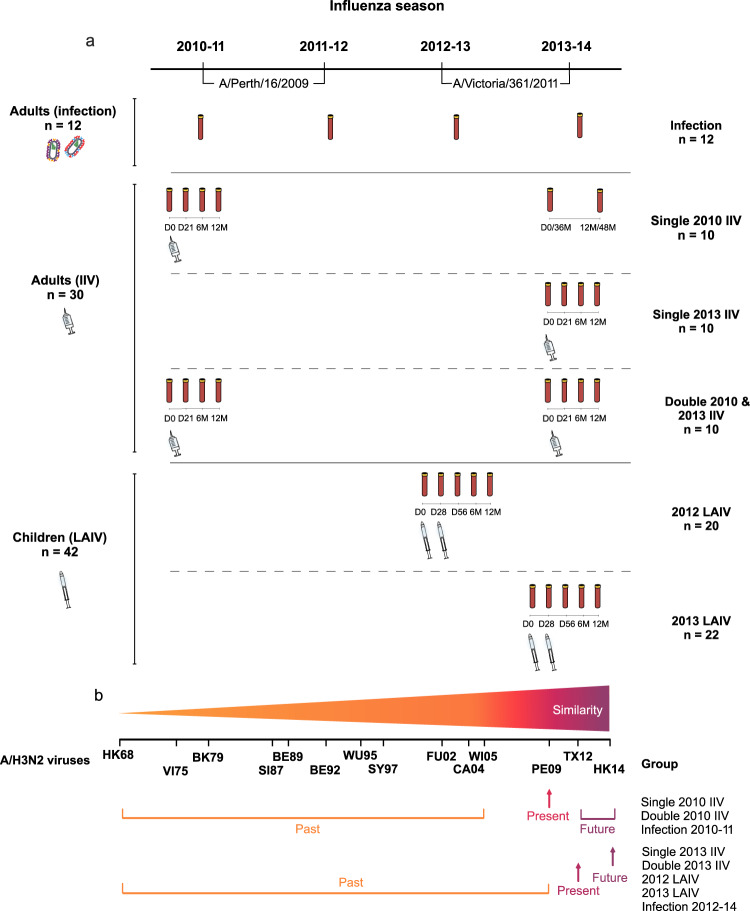
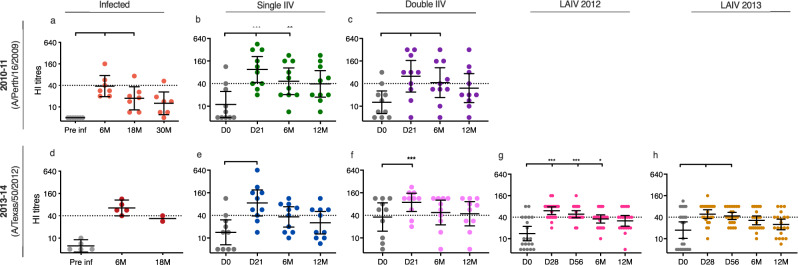
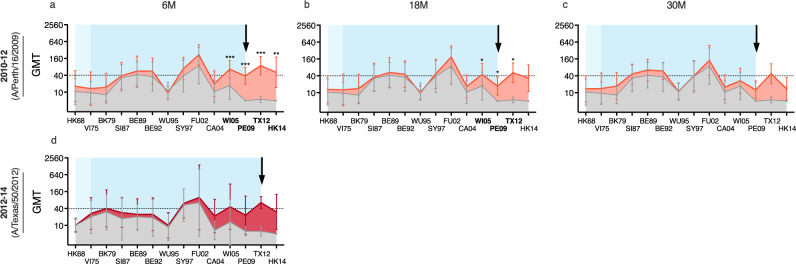
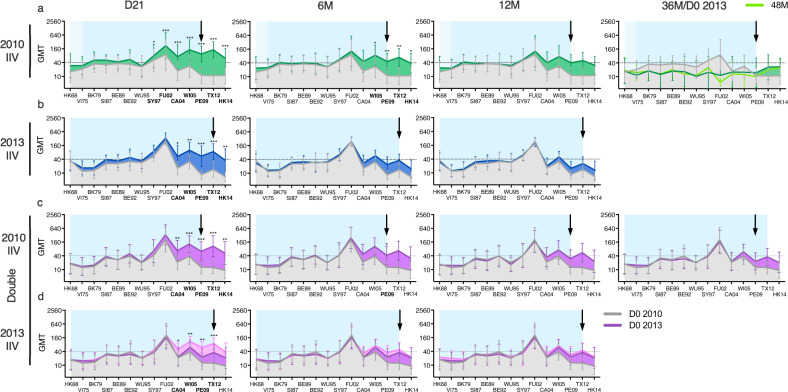
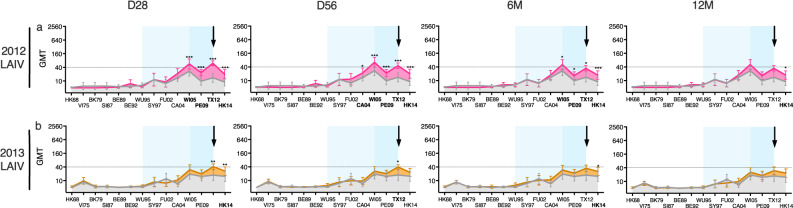
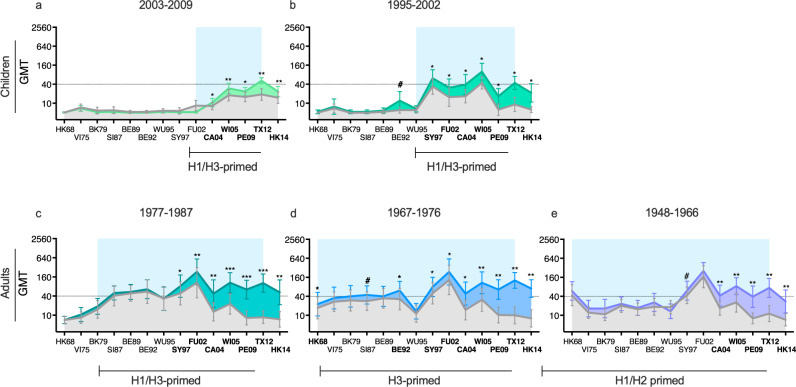
History of influenza A/H3N2 exposure, especially childhood infection, shape antibody responses after influenza vaccination and infection, but have not been extensively studied. We investigated the breadth and durability of influenza A/H3N2-specific hemagglutinin-inhibition antibodies after live-attenuated influenza vaccine in children (aged 3-17 years, n = 42), and after inactivated influenza vaccine or infection in adults (aged 22-61 years, n = 42) using 14 antigenically distinct A/H3N2 viruses circulating from 1968 to 2018. We found that vaccination and infection elicited cross-reactive antibody responses, predominantly directed against newer or future strains. Childhood H3-priming increased the breadth and magnitude of back-boosted A/H3N2-specific antibodies in adults. Broader and more durable A/H3N2-specific antibodies were observed in repeatedly vaccinated adults than in children and previously unvaccinated adults. Our findings suggest that early A/H3N2 exposure and frequent seasonal vaccination could increase the breadth and seropositivity of antibody responses, which may improve vaccine protection against future viruses.
© 2022. The Author(s).
Conflict of interest statement
All authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures

References
-
- World Health Organization. Influenza (Seasonal), https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (2018).
LinkOut - more resources
Full Text Sources
