

Metabolic Engineering of Bacillus amyloliquefaciens to Efficiently Synthesize L-Ornithine From Inulin
- PMID: 35757793
- PMCID: PMC9214239
- DOI: 10.3389/fbioe.2022.905110
Metabolic Engineering of Bacillus amyloliquefaciens to Efficiently Synthesize L-Ornithine From Inulin
Abstract
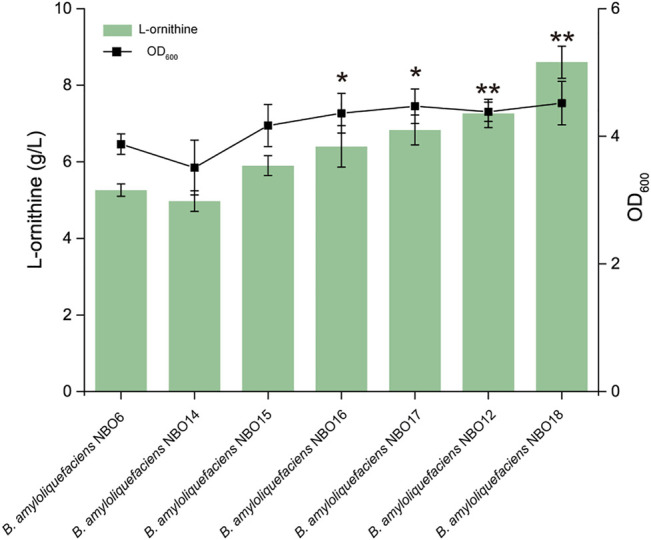
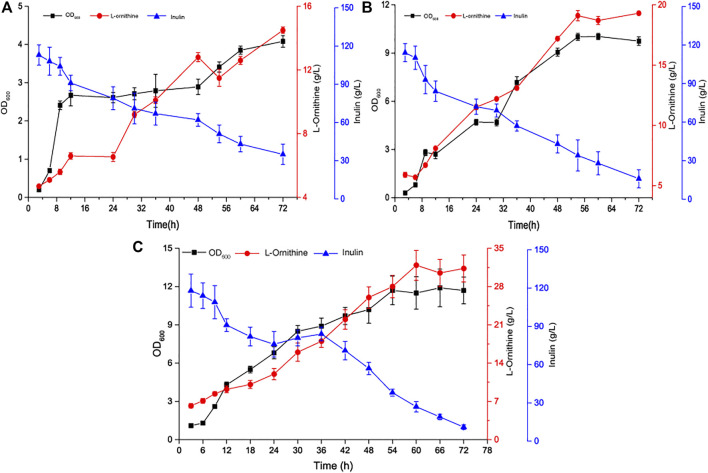
Bacillus amyloliquefaciens is the dominant strain used to produce γ-polyglutamic acid from inulin, a non-grain raw material. B. amyloliquefaciens has a highly efficient tricarboxylic acid cycle metabolic flux and glutamate synthesis ability. These features confer great potential for the synthesis of glutamate derivatives. However, it is challenging to efficiently convert high levels of glutamate to a particular glutamate derivative. Here, we conducted a systematic study on the biosynthesis of L-ornithine by B. amyloliquefaciens using inulin. First, the polyglutamate synthase gene pgsBCA of B. amyloliquefaciens NB was knocked out to hinder polyglutamate synthesis, resulting in the accumulation of intracellular glutamate and ATP. Second, a modular engineering strategy was applied to coordinate the degradation pathway, precursor competition pathway, and L-ornithine synthesis pathway to prompt high levels of intracellular precursor glutamate for l-ornithine synthesis. In addition, the high-efficiency L-ornithine transporter was further screened and overexpressed to reduce the feedback inhibition of L-ornithine on the synthesis pathway. Combining these strategies with further fermentation optimizations, we achieved a final L-ornithine titer of 31.3 g/L from inulin. Overall, these strategies hold great potential for strengthening microbial synthesis of high value-added products derived from glutamate.
Keywords: Bacillus amyloliquefaciens; L-Ornithine; L-ornithine transporter; modular metabolic engineering; non-grain raw materials.
Copyright © 2022 Zhu, Hu, Yan, Du, Pan, Li, Xu and Luo.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



References
LinkOut - more resources
Full Text Sources