

Microdomain Protein Nce102 Is a Local Sensor of Plasma Membrane Sphingolipid Balance
- PMID: 35758748
- PMCID: PMC9431316
- DOI: 10.1128/spectrum.01961-22
Microdomain Protein Nce102 Is a Local Sensor of Plasma Membrane Sphingolipid Balance
Abstract
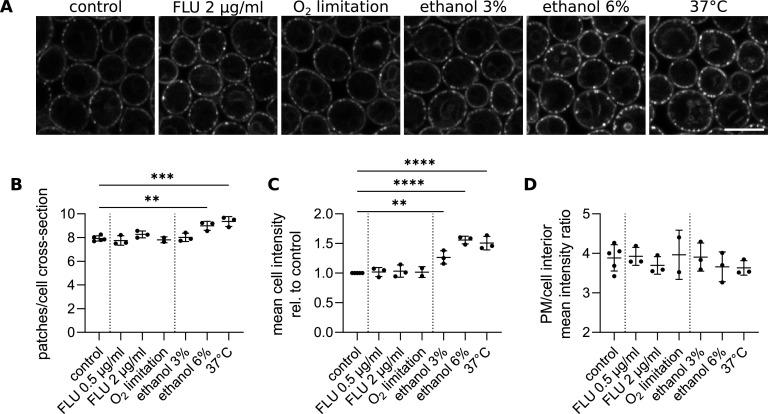
Sphingolipids are essential building blocks of eukaryotic membranes and important signaling molecules that are regulated tightly in response to environmental and physiological inputs. While their biosynthetic pathway has been well-described, the mechanisms that facilitate the perception of sphingolipid levels at the plasma membrane remain to be uncovered. In Saccharomyces cerevisiae, the Nce102 protein has been proposed to function as a sphingolipid sensor as it changes its plasma membrane distribution in response to sphingolipid biosynthesis inhibition. We show that Nce102 redistributes specifically in regions of increased sphingolipid demand, e.g., membranes of nascent buds. Furthermore, we report that the production of Nce102 increases following sphingolipid biosynthesis inhibition and that Nce102 is internalized when excess sphingolipid precursors are supplied. This finding suggests that the total amount of Nce102 in the plasma membrane is a measure of the current need for sphingolipids, whereas its local distribution marks sites of high sphingolipid demand. The physiological role of Nce102 in the regulation of sphingolipid synthesis is demonstrated by mass spectrometry analysis showing reduced levels of hydroxylated complex sphingolipids in response to heat stress in the nce102Δ deletion mutant. We also demonstrate that Nce102 behaves analogously in the widespread human fungal pathogen Candida albicans, suggesting a conserved principle of local sphingolipid control across species. IMPORTANCE Microorganisms are challenged constantly by their rapidly changing environment. To survive, they have developed diverse mechanisms to quickly perceive stressful situations and adapt to them appropriately. The primary site of both stress sensing and adaptation is the plasma membrane. We identified the yeast protein Nce102 as a marker of local sphingolipid levels and fluidity in the plasma membrane. Nce102 is an important structural and functional component of the membrane compartment Can1 (MCC), a plasma membrane microdomain stabilized by a large cytosolic hemitubular protein scaffold, the eisosome. The MCC/eisosomes are widely conserved among fungi and unicellular algae. To determine if Nce102 carries out similar functions in other organisms, we analyzed the human fungal pathogen Candida albicans and found that Nce102 responds to sphingolipid levels also in this organism, which has potential applications for the development of novel therapeutic approaches. The presented study represents a valuable model for how organisms regulate plasma membrane sphingolipids.
Keywords: eisosome; microdomain; plasma membrane; sphingolipid; stress sensor.
Conflict of interest statement
The authors declare a conflict of interest. We declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. M.D.P. is a co-founder and Chief Scientific Officer (CSO) of MicroRid Technologies Inc.
Figures







References
-
- Young ME, Karpova TS, Brugger B, Moschenross DM, Wang GK, Schneiter R, Wieland FT, Cooper JA. 2002. The Sur7p family defines novel cortical domains in Saccharomyces cerevisiae, affects sphingolipid metabolism, and is involved in sporulation. Mol Cell Biol 22:927–934. doi: 10.1128/MCB.22.3.927-934.2002. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
