

Long-distance facilitation of coastal ecosystem structure and resilience
- PMID: 35759652
- PMCID: PMC9282383
- DOI: 10.1073/pnas.2123274119
Long-distance facilitation of coastal ecosystem structure and resilience
Abstract
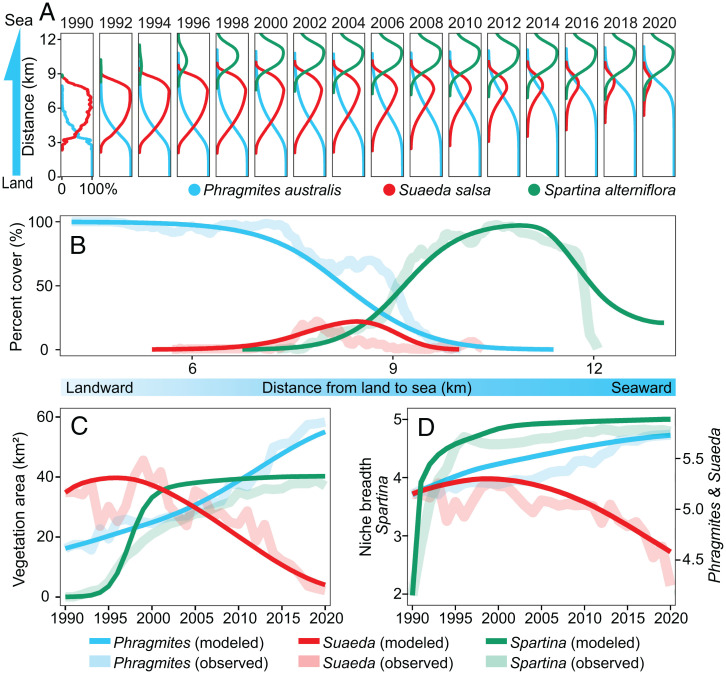
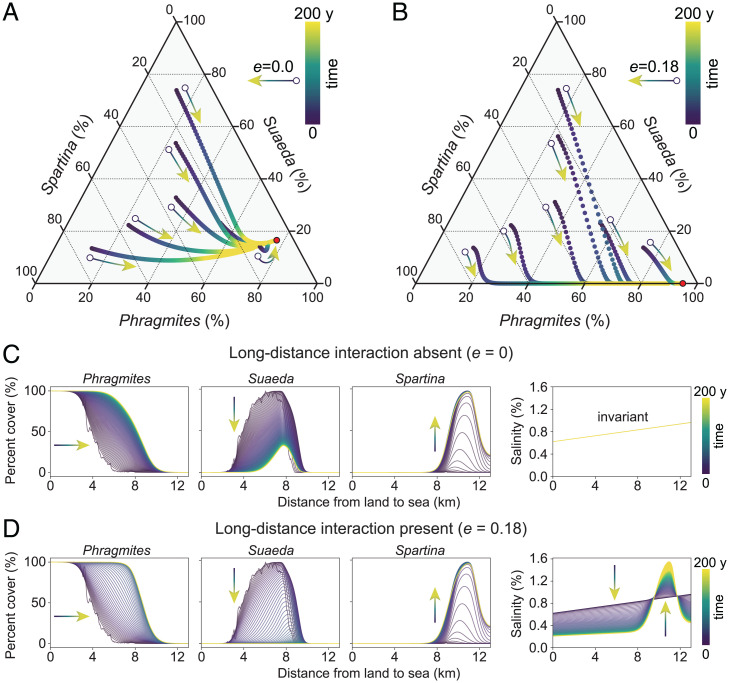
Biotic interactions that hierarchically organize ecosystems by driving ecological and evolutionary processes across spatial scales are ubiquitous in our biosphere. Biotic interactions have been extensively studied at local and global scales, but how long-distance, cross-ecosystem interactions at intermediate landscape scales influence the structure, function, and resilience of ecological systems remains poorly understood. We used remote sensing, modeling, and field data to test the hypothesis that the long-distance impact of an invasive species dramatically affects one of the largest tidal flat ecosystems in East Asia. We found that the invasion of exotic cordgrass Spartina alterniflora can produce long-distance effects on native species up to 10 km away, driving decadal coastal ecosystem transitions. The invasive cordgrass at low elevations facilitated the expansion of the native reed Phragmites australis at high elevations, leading to the massive loss and reduced resilience of the iconic Suaeda salsa "Red Beach" marshes at intermediate elevations, largely as a consequence of reduced soil salinity across the landscape. Our results illustrate the complex role that long-distance interactions can play in shaping landscape structure and ecosystem resilience and in bridging the gap between local and global biotic interactions.
Keywords: biological invasion; biotic interaction; coastal saltmarsh; long-distance interaction; resilience.
Conflict of interest statement
The authors declare no competing interest.
Figures




References
-
- Hutchinson G. E., Homage to Santa Rosalia or why are there so many kinds of animals? Am. Nat. 93, 145–159 (1959).
-
- Andresen E., Arroyo-Rodríguez V., Escobar F., “Tropical biodiversity: The importance of biotic interactions for its origin, maintenance, function, and conservation” in Ecological Networks in the Tropics, Dáttilo W., Rico-Gray V., Eds. (Springer, 2018), pp. 1–13.
-
- Brown J. H., Two decades of homage to Santa Rosalia: Toward a general theory of diversity. Am. Zool. 21, 877–888 (1981).
-
- Vellend M., The Theory of Ecological Communities (MPB-57) (Princeton University Press, 2016).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
