

Brown adipose tissue-derived MaR2 contributes to cold-induced resolution of inflammation
- PMID: 35760872
- PMCID: PMC9792164
- DOI: 10.1038/s42255-022-00590-0
Brown adipose tissue-derived MaR2 contributes to cold-induced resolution of inflammation
Abstract
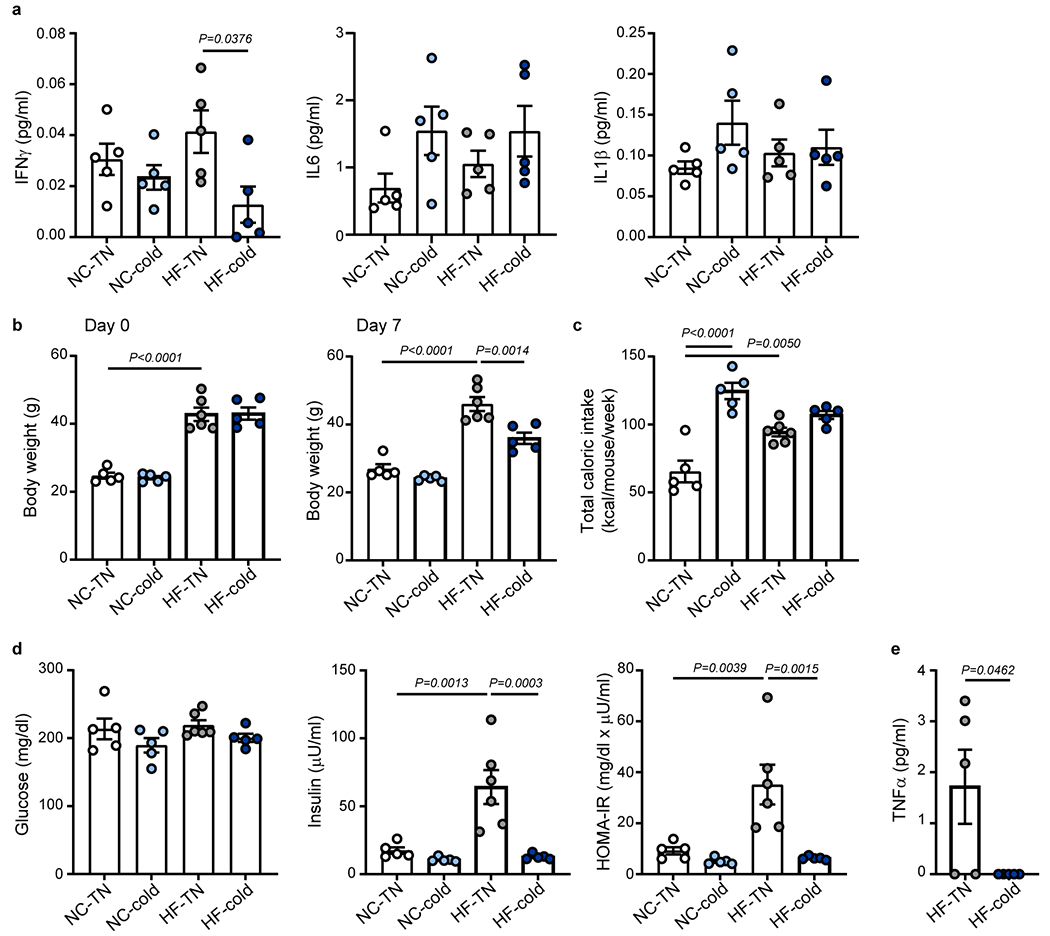
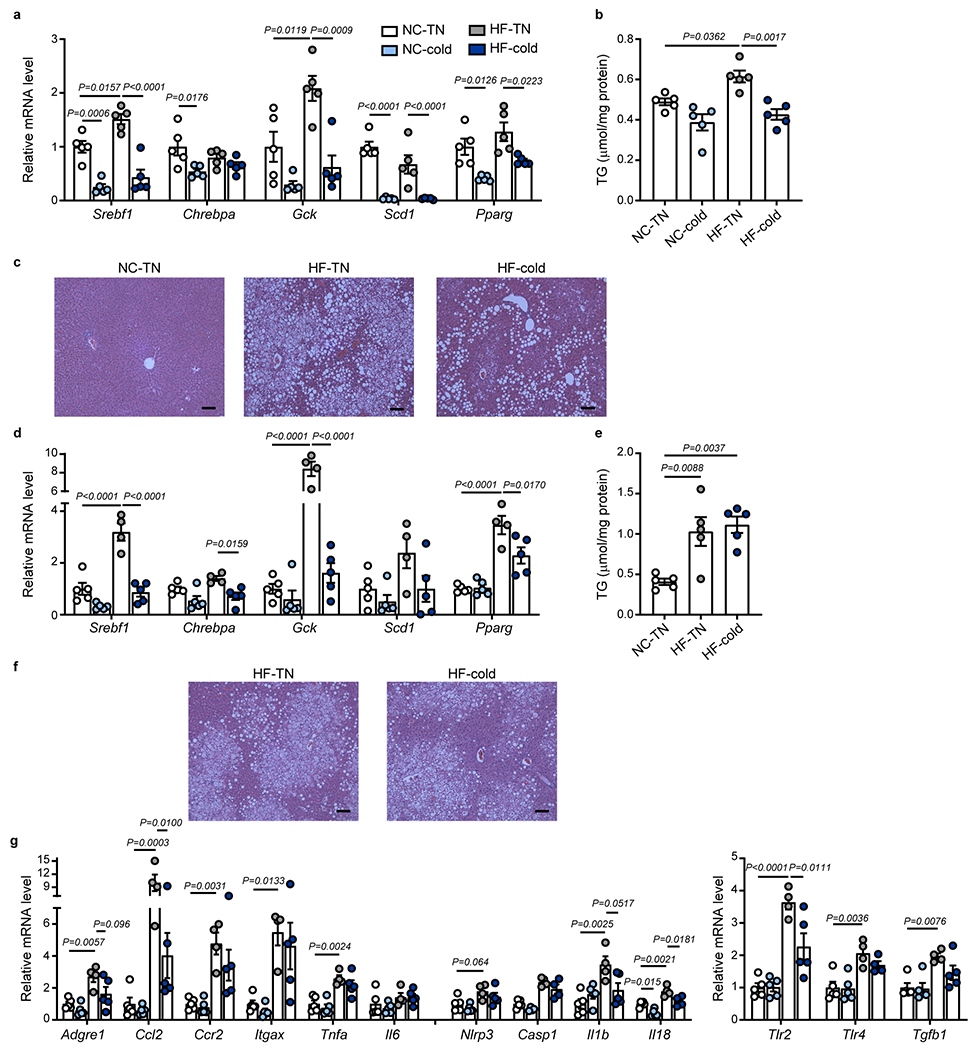
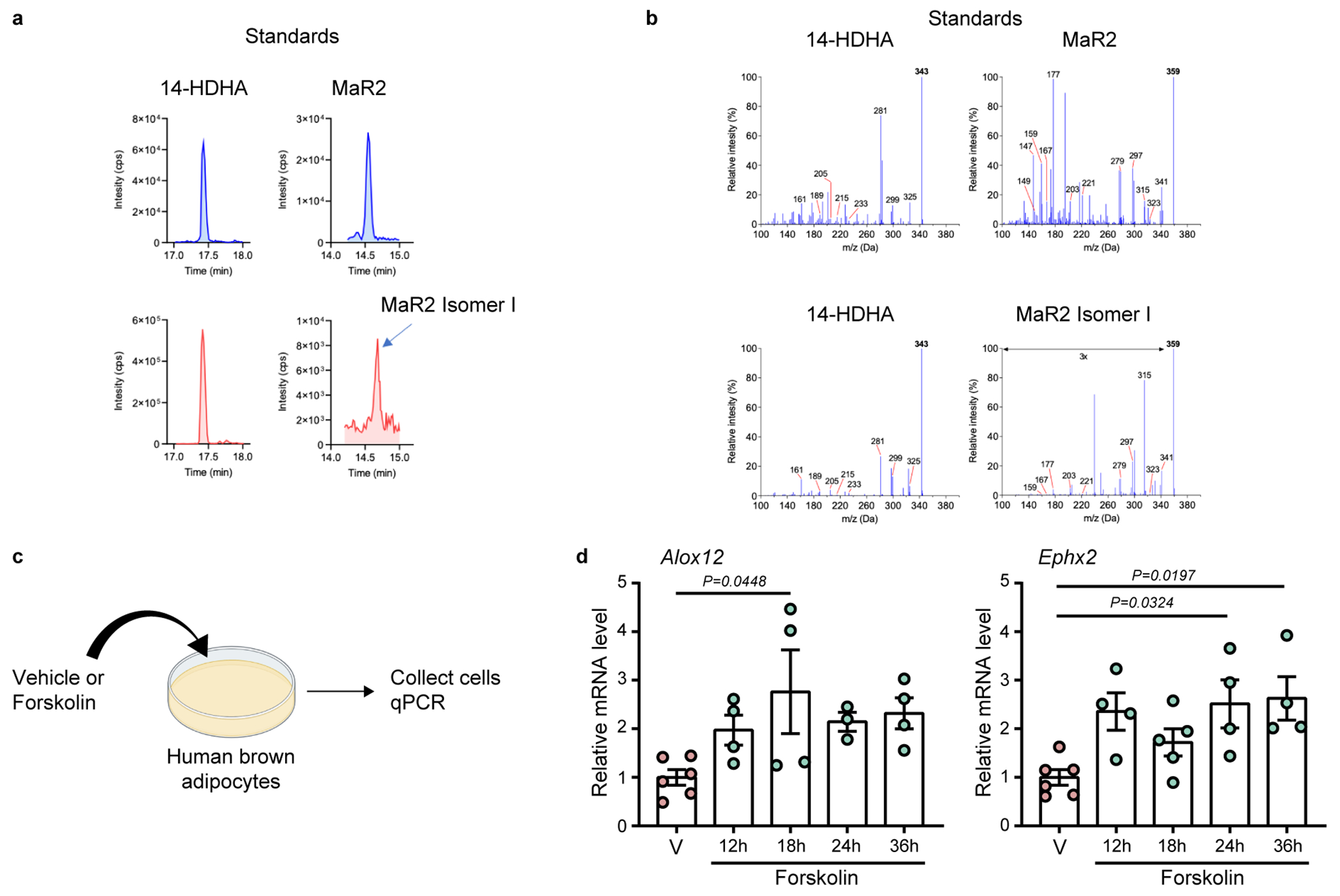
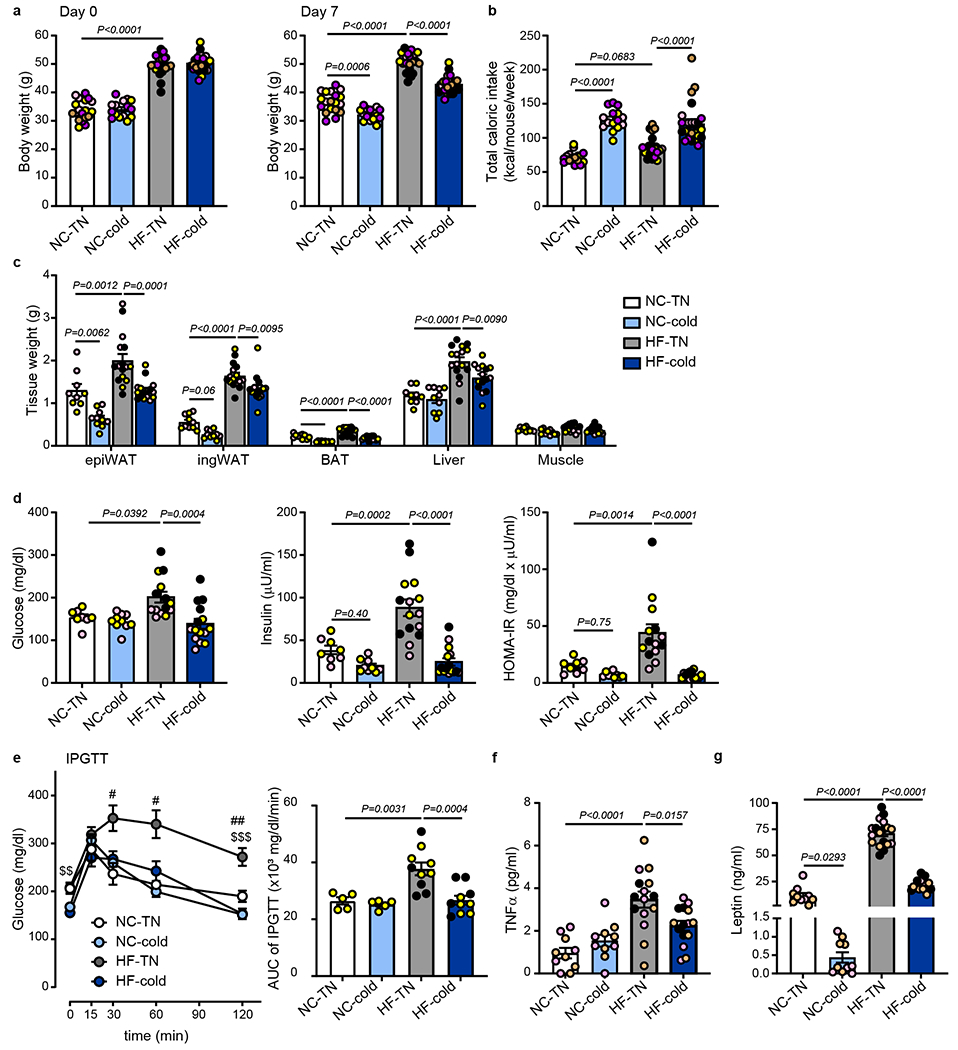
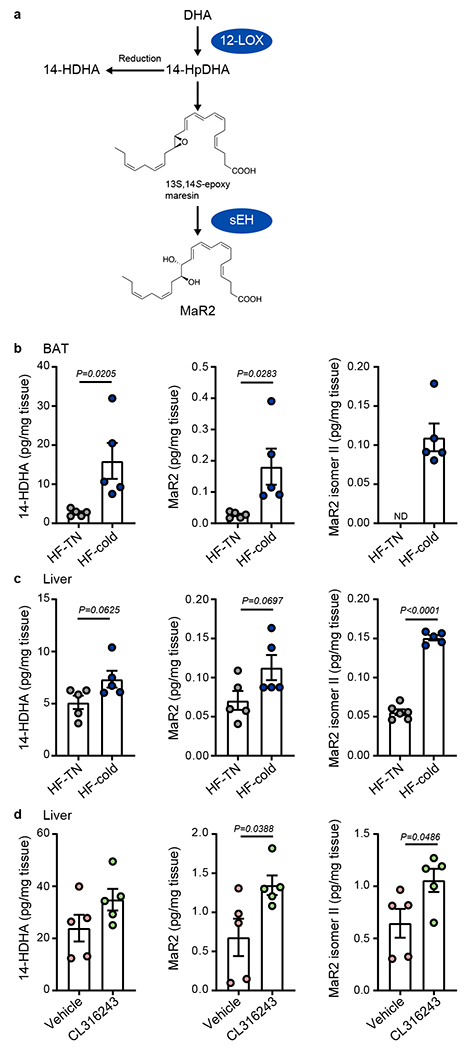
Obesity induces chronic inflammation resulting in insulin resistance and metabolic disorders. Cold exposure can improve insulin sensitivity in humans and rodents, but the mechanisms have not been fully elucidated. Here, we find that cold resolves obesity-induced inflammation and insulin resistance and improves glucose tolerance in diet-induced obese mice. The beneficial effects of cold exposure on improving obesity-induced inflammation and insulin resistance depend on brown adipose tissue (BAT) and liver. Using targeted liquid chromatography with tandem mass spectrometry, we discovered that cold and β3-adrenergic stimulation promote BAT to produce maresin 2 (MaR2), a member of the specialized pro-resolving mediators of bioactive lipids that play a role in the resolution of inflammation. Notably, MaR2 reduces inflammation in obesity in part by targeting macrophages in the liver. Thus, BAT-derived MaR2 could contribute to the beneficial effects of BAT activation in resolving obesity-induced inflammation and may inform therapeutic approaches to combat obesity and its complications.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing Interests Statement
M.S. and Y.-H.T. are inventors of a pending provisional patent application related to maresin 2 and metabolic therapeutics.
Figures












Comment in
-
Brown fat resolves hepatic inflammation in obesity.Nat Metab. 2022 Jun;4(6):649-650. doi: 10.1038/s42255-022-00596-8. Nat Metab. 2022. PMID: 35760870 No abstract available.
References
-
- Blüher M Obesity: global epidemiology and pathogenesis. Nature reviews. Endocrinology 15, 288–298 (2019). - PubMed
-
- Esser N, Legrand-Poels S, Piette J, Scheen AJ & Paquot N Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes research and clinical practice 105, 141–150 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
