

Maternal gut microbiota Bifidobacterium promotes placental morphogenesis, nutrient transport and fetal growth in mice
- PMID: 35760917
- PMCID: PMC9236968
- DOI: 10.1007/s00018-022-04379-y
Maternal gut microbiota Bifidobacterium promotes placental morphogenesis, nutrient transport and fetal growth in mice
Abstract
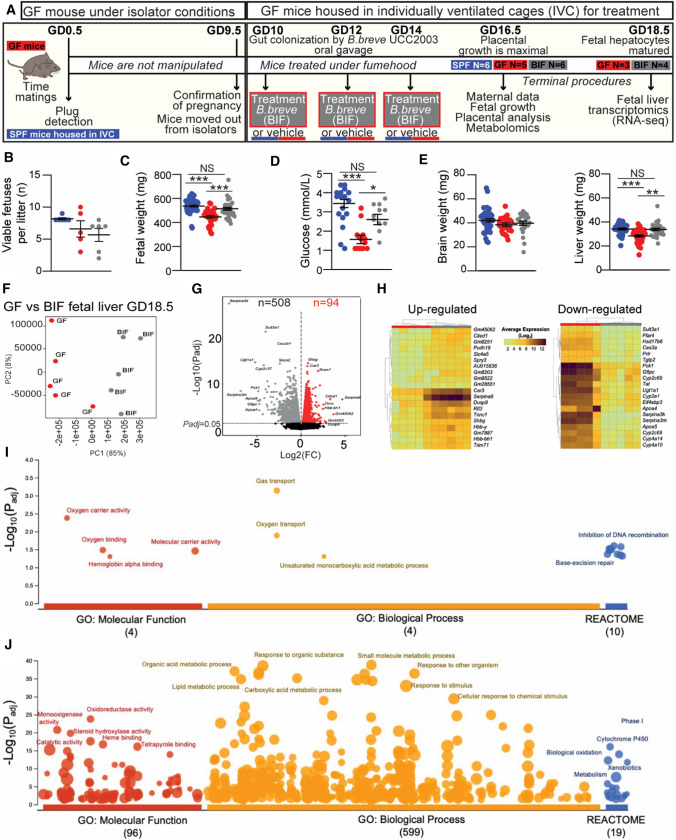
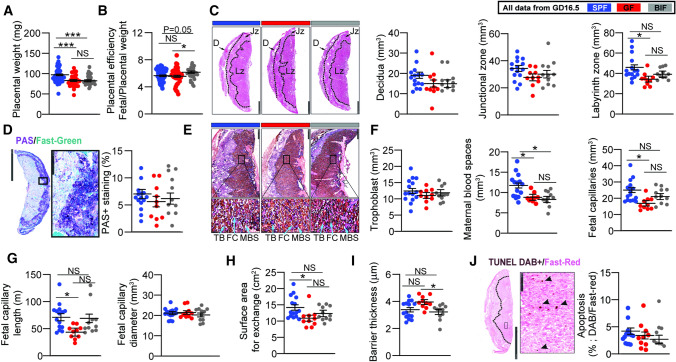
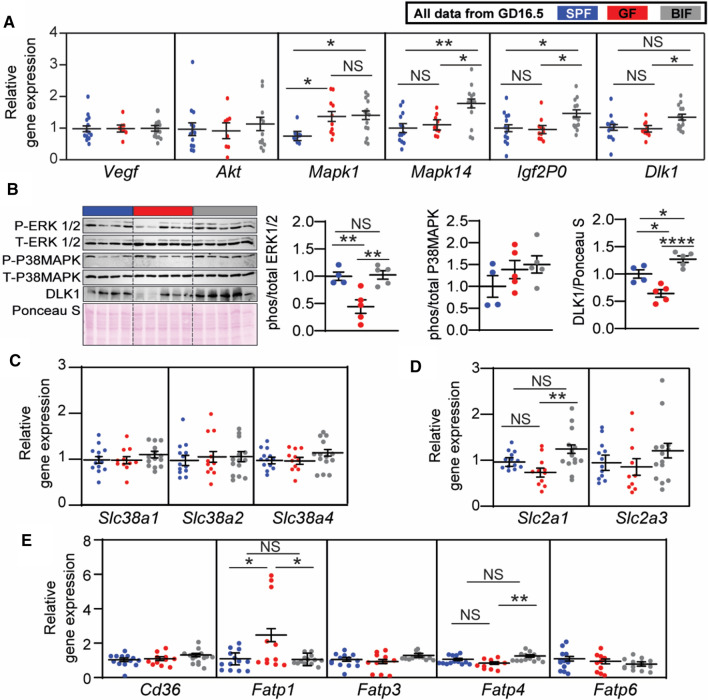
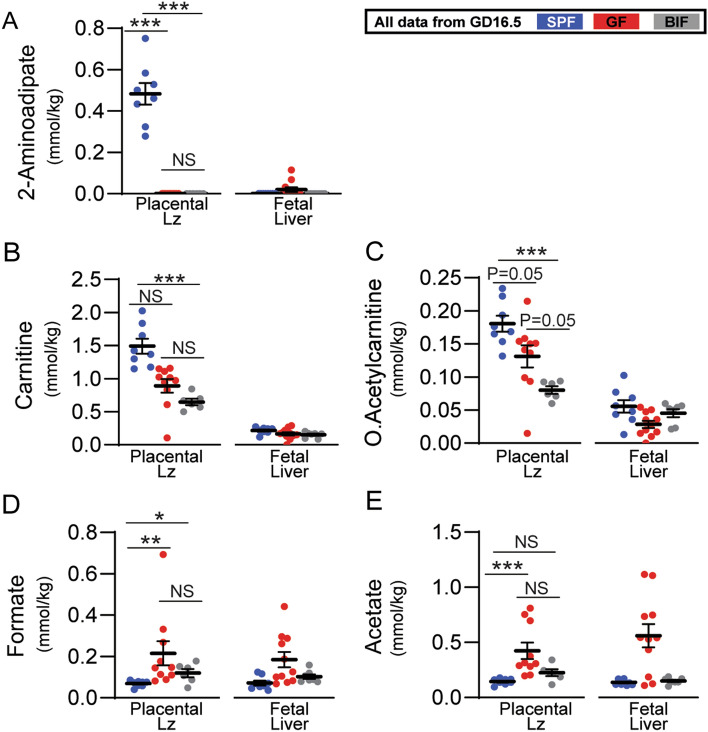
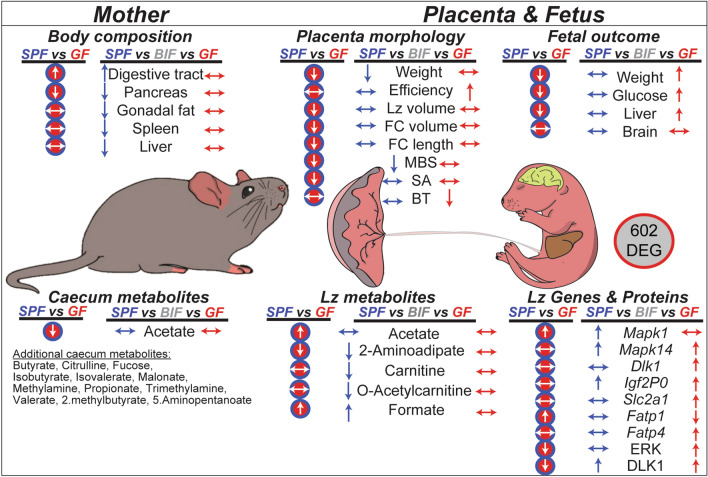
The gut microbiota plays a central role in regulating host metabolism. While substantial progress has been made in discerning how the microbiota influences host functions post birth and beyond, little is known about how key members of the maternal gut microbiota can influence feto-placental growth. Notably, in pregnant women, Bifidobacterium represents a key beneficial microbiota genus, with levels observed to increase across pregnancy. Here, using germ-free and specific-pathogen-free mice, we demonstrate that the bacterium Bifidobacterium breve UCC2003 modulates maternal body adaptations, placental structure and nutrient transporter capacity, with implications for fetal metabolism and growth. Maternal and placental metabolome were affected by maternal gut microbiota (i.e. acetate, formate and carnitine). Histological analysis of the placenta confirmed that Bifidobacterium modifies placental structure via changes in Igf2P0, Dlk1, Mapk1 and Mapk14 expression. Additionally, B. breve UCC2003, acting through Slc2a1 and Fatp1-4 transporters, was shown to restore fetal glycaemia and fetal growth in association with changes in the fetal hepatic transcriptome. Our work emphasizes the importance of the maternal gut microbiota on feto-placental development and sets a foundation for future research towards the use of probiotics during pregnancy.
Keywords: Bifidobacterium; Fetus; Metabolism; Microbiota; Pregnancy.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
MeSH terms
Grants and funding
- MR/R022690/1/MRC_/Medical Research Council/United Kingdom
- BB/R012490/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/F/000PR10356/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/F/000PR10353/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 220456/Z/20/Z/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Miscellaneous
