

Identification of Pathogenic Immune Cell Subsets Associated With Checkpoint Inhibitor-Induced Myocarditis
- PMID: 35762356
- PMCID: PMC9397491
- DOI: 10.1161/CIRCULATIONAHA.121.056730
Identification of Pathogenic Immune Cell Subsets Associated With Checkpoint Inhibitor-Induced Myocarditis
Abstract
Background: Immune checkpoint inhibitors (ICIs) are monoclonal antibodies used to activate the immune system against tumor cells. Despite therapeutic benefits, ICIs have the potential to cause immune-related adverse events such as myocarditis, a rare but serious side effect with up to 50% mortality in affected patients. Histologically, patients with ICI myocarditis have lymphocytic infiltrates in the heart, implicating T cell-mediated mechanisms. However, the precise pathological immune subsets and molecular changes in ICI myocarditis are unknown.
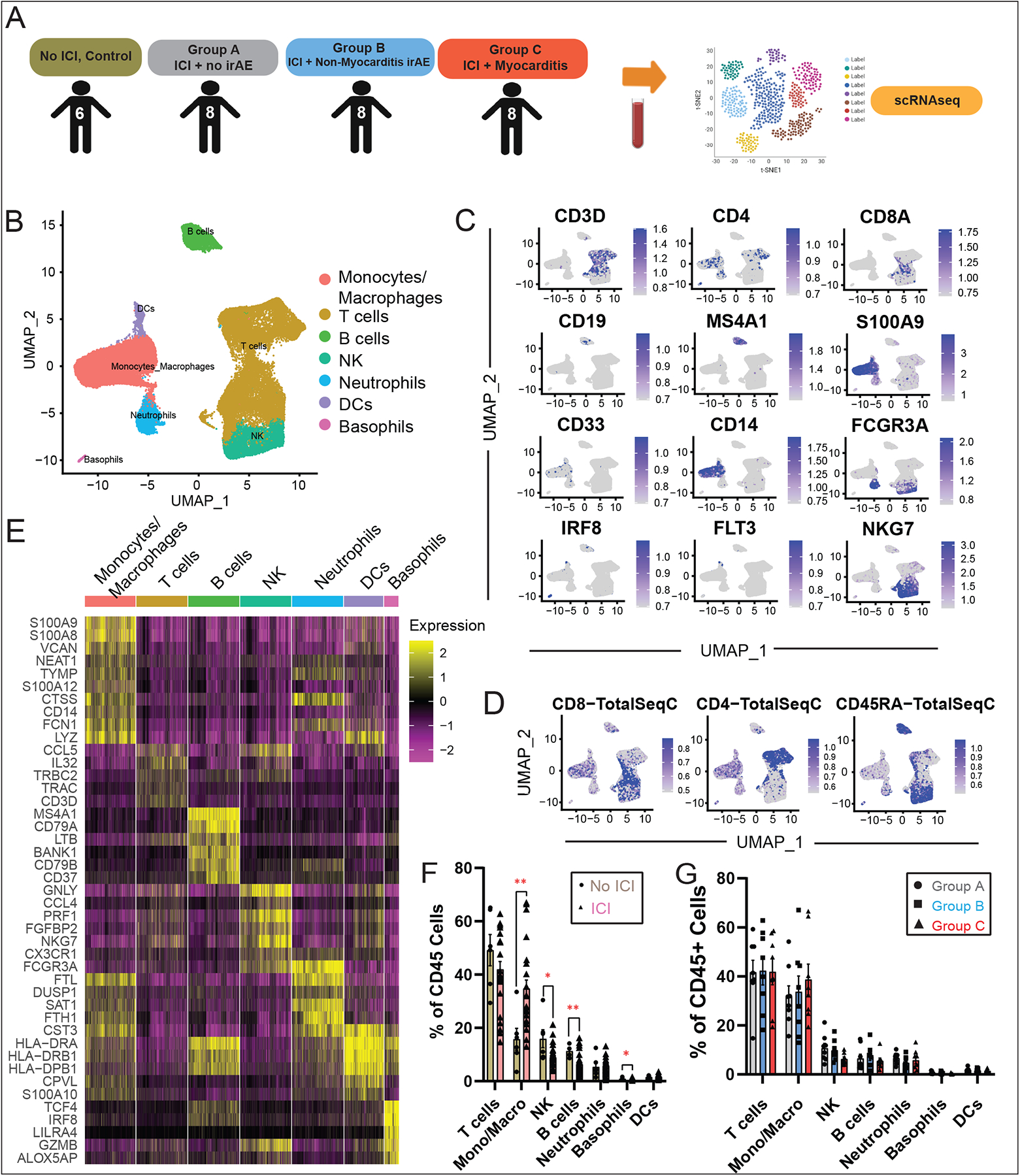
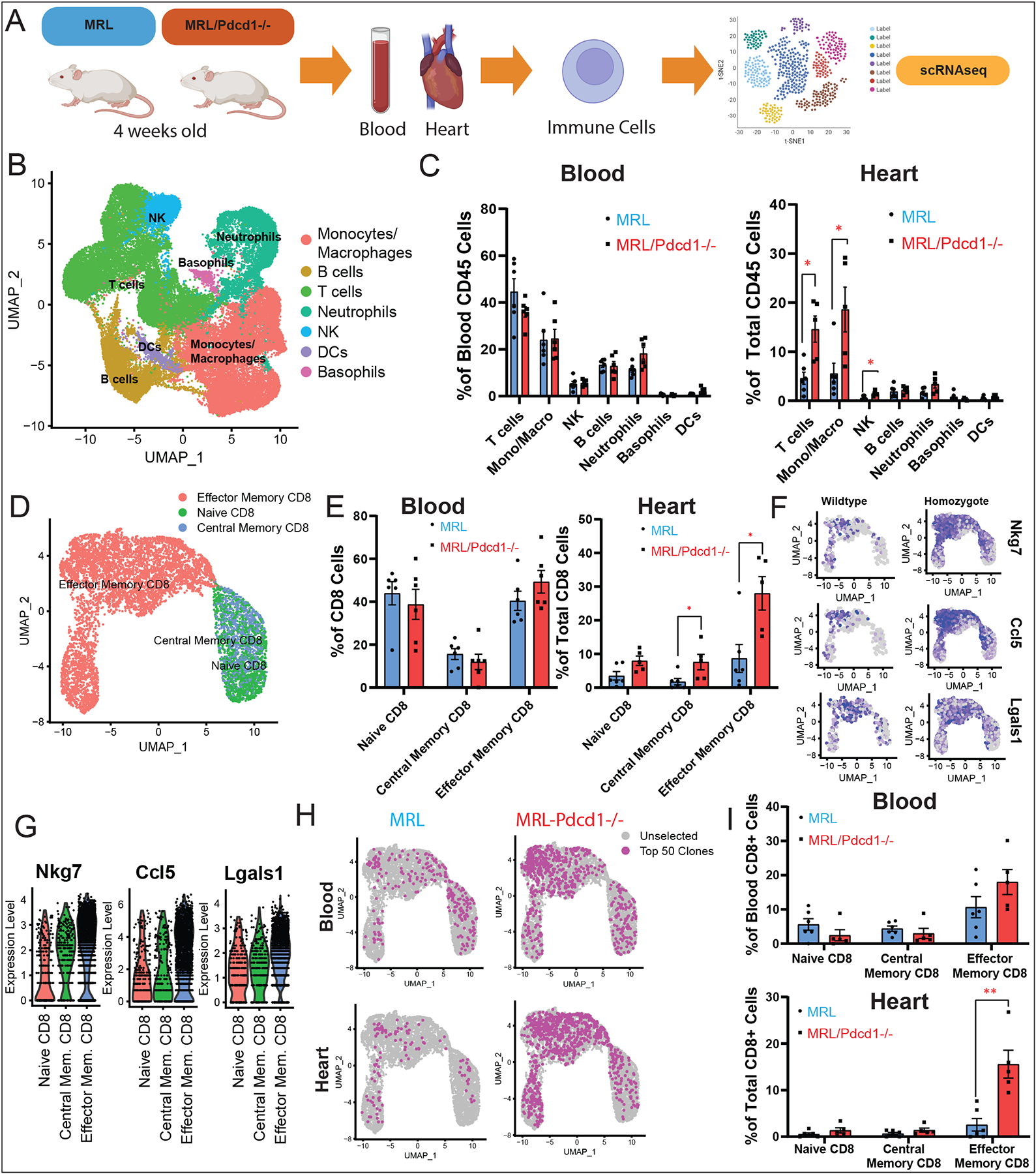
Methods: To identify immune subset(s) associated with ICI myocarditis, we performed time-of-flight mass cytometry on peripheral blood mononuclear cells from 52 individuals: 29 patients with autoimmune adverse events (immune-related adverse events) on ICI, including 8 patients with ICI myocarditis, and 23 healthy control subjects. We also used multiomics single-cell technology to immunophenotype 30 patients/control subjects using single-cell RNA sequencing, single-cell T-cell receptor sequencing, and cellular indexing of transcriptomes and epitopes by sequencing with feature barcoding for surface marker expression confirmation. To correlate between the blood and the heart, we performed single-cell RNA sequencing/T-cell receptor sequencing/cellular indexing of transcriptomes and epitopes by sequencing on MRL/Pdcd1-/- (Murphy Roths large/programmed death-1-deficient) mice with spontaneous myocarditis.
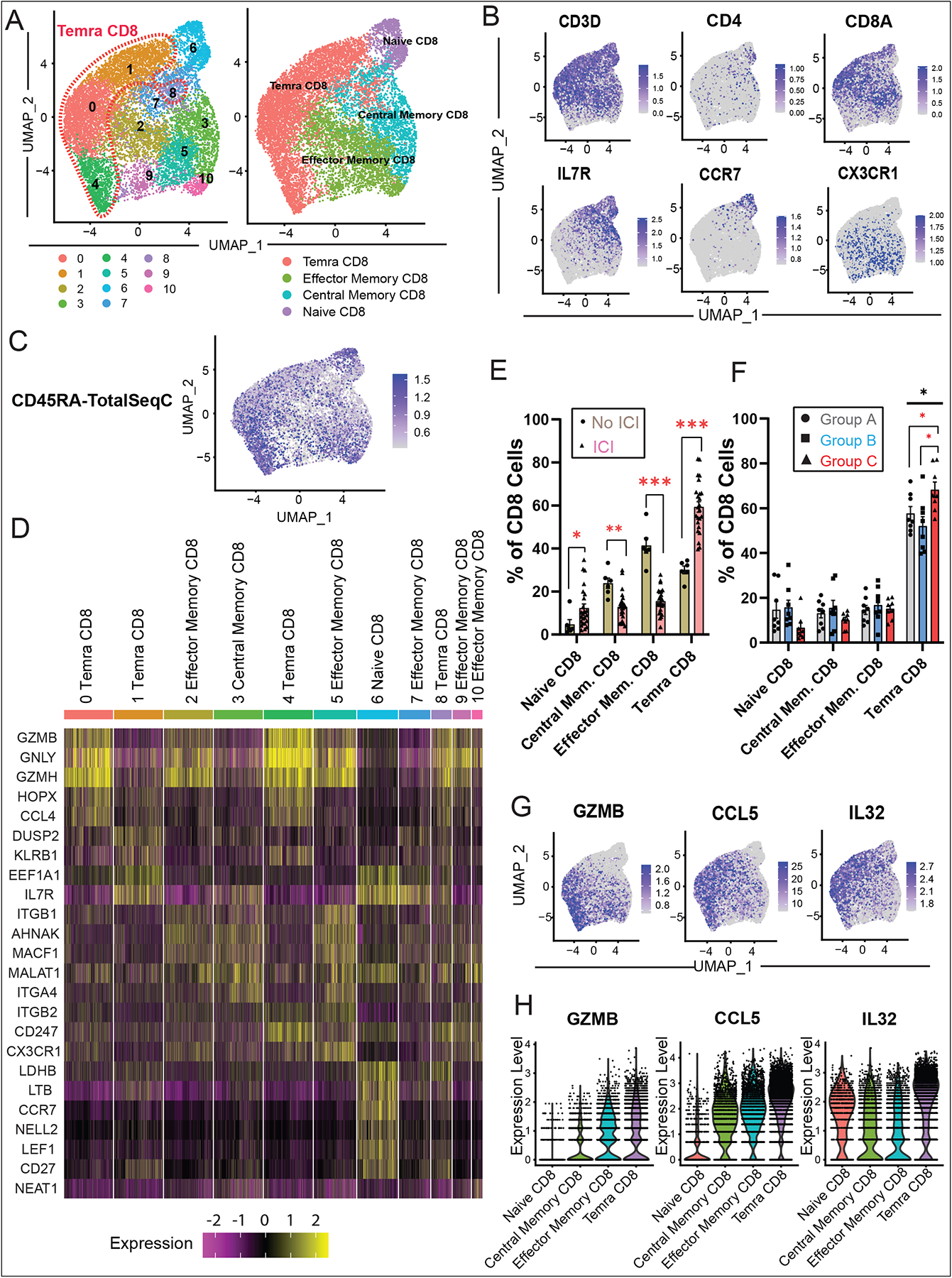
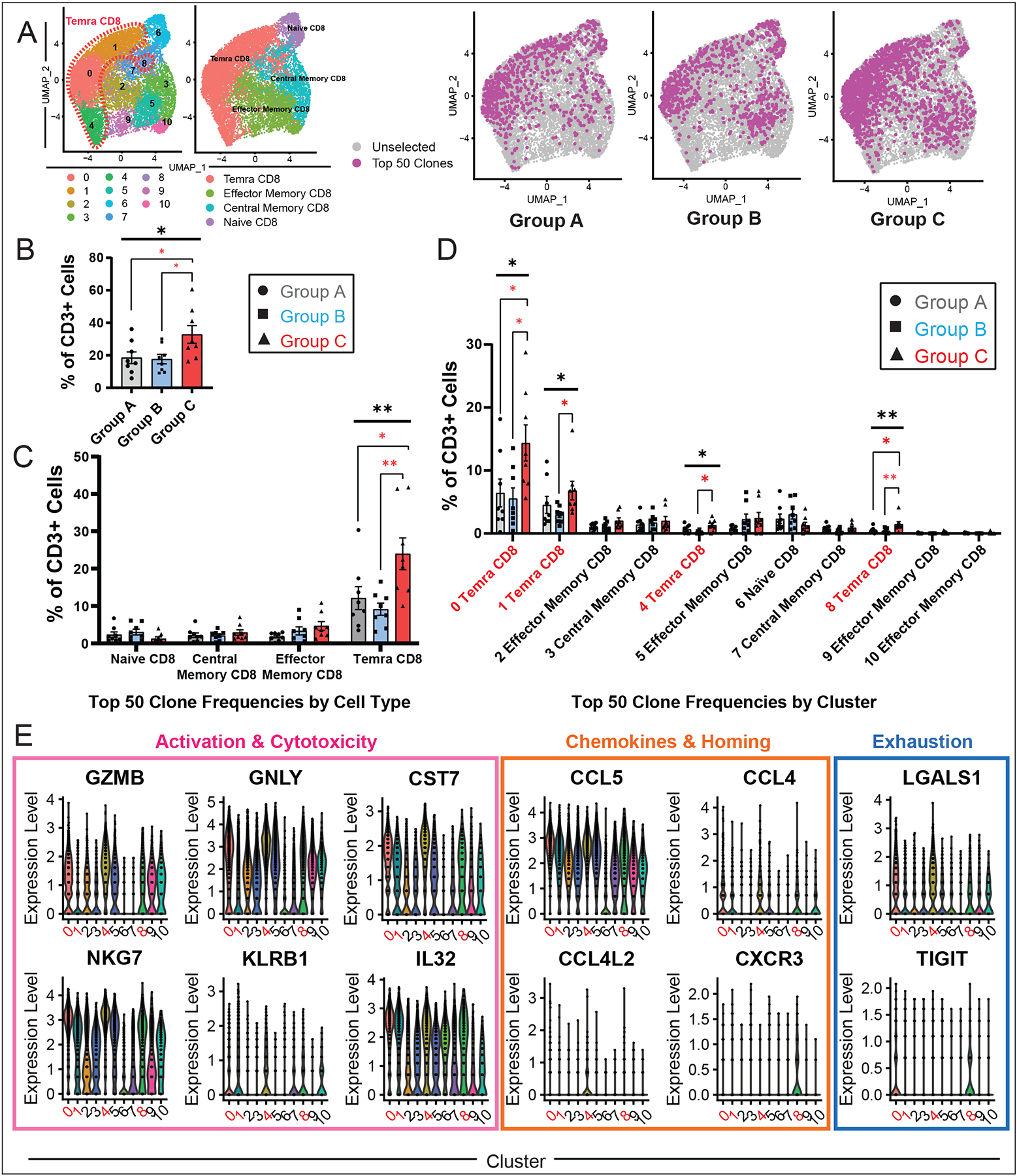
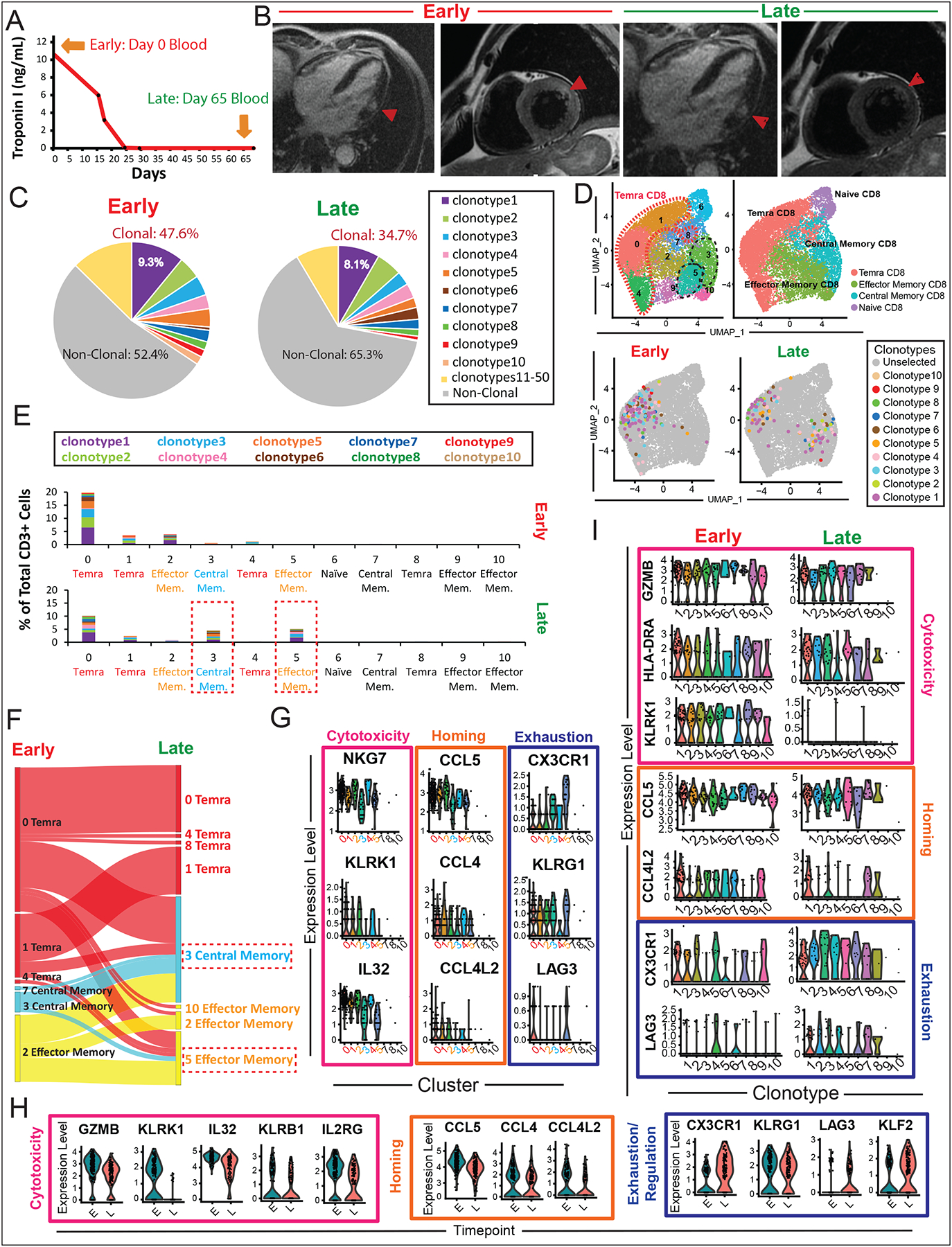
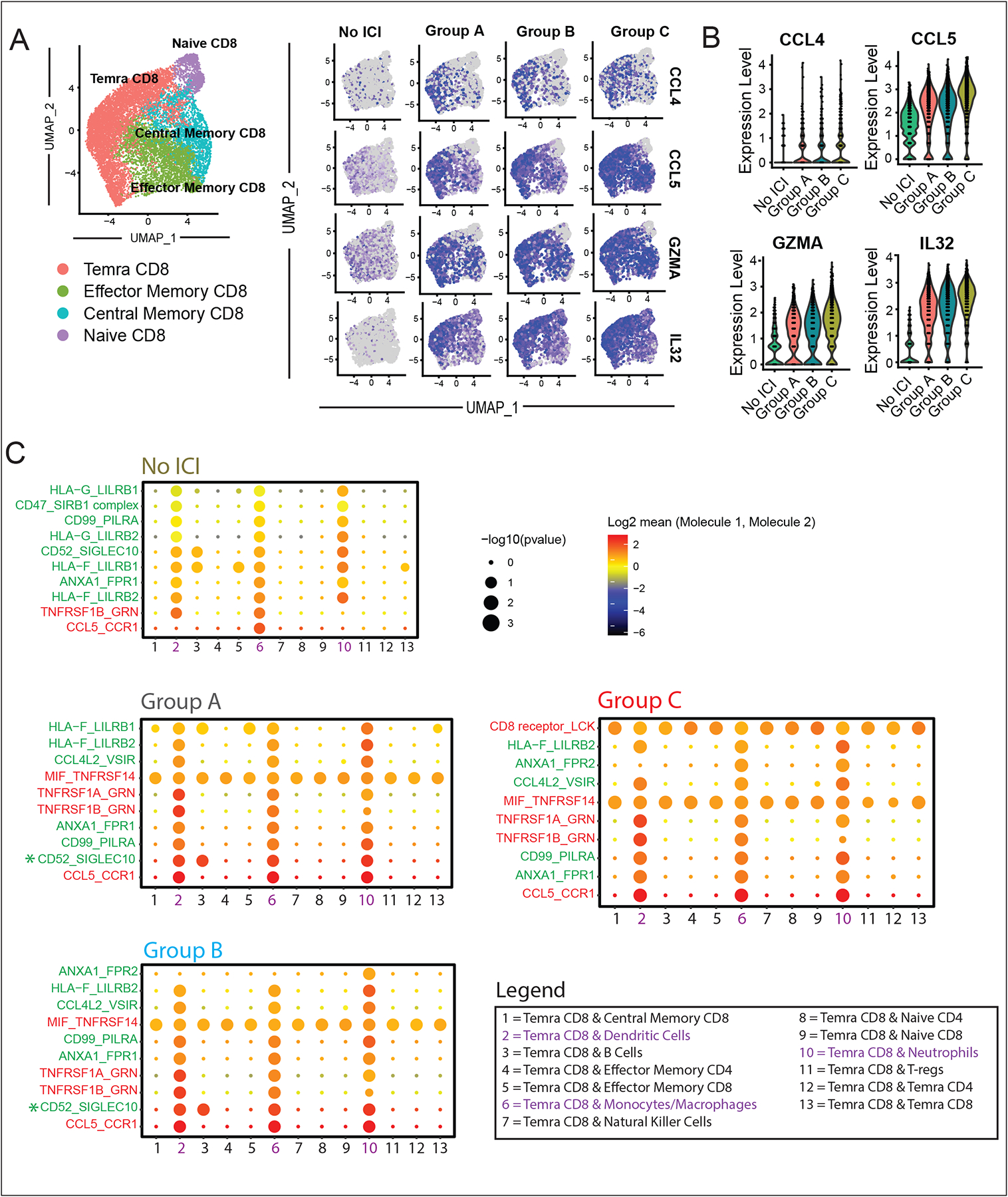
Results: Using these complementary approaches, we found an expansion of cytotoxic CD8+ T effector cells re-expressing CD45RA (Temra CD8+ cells) in patients with ICI myocarditis compared with control subjects. T-cell receptor sequencing demonstrated that these CD8+ Temra cells were clonally expanded in patients with myocarditis compared with control subjects. Transcriptomic analysis of these Temra CD8+ clones confirmed a highly activated and cytotoxic phenotype. Longitudinal study demonstrated progression of these Temra CD8+ cells into an exhausted phenotype 2 months after treatment with glucocorticoids. Differential expression analysis demonstrated elevated expression levels of proinflammatory chemokines (CCL5/CCL4/CCL4L2) in the clonally expanded Temra CD8+ cells, and ligand receptor analysis demonstrated their interactions with innate immune cells, including monocytes/macrophages, dendritic cells, and neutrophils, as well as the absence of key anti-inflammatory signals. To complement the human study, we performed single-cell RNA sequencing/T-cell receptor sequencing/cellular indexing of transcriptomes and epitopes by sequencing in Pdcd1-/- mice with spontaneous myocarditis and found analogous expansions of cytotoxic clonal effector CD8+ cells in both blood and hearts of such mice compared with controls.
Conclusions: Clonal cytotoxic Temra CD8+ cells are significantly increased in the blood of patients with ICI myocarditis, corresponding to an analogous increase in effector cytotoxic CD8+ cells in the blood/hearts of Pdcd1-/- mice with myocarditis. These expanded effector CD8+ cells have unique transcriptional changes, including upregulation of chemokines CCL5/CCL4/CCL4L2, which may serve as attractive diagnostic/therapeutic targets for reducing life-threatening cardiac immune-related adverse events in ICI-treated patients with cancer.
Keywords: immune checkpoint inhibitors; immunophenotyping; immunotherapy; myocarditis; single-cell analysis.
Figures

Comment in
-
An Immune Checkpoint Inhibitor Heart: How CD45RA+ Effector Memory CD8+ T Cells (Temra) Are Implicated in Immune Checkpoint Inhibitor Myocarditis.Circulation. 2022 Jul 26;146(4):336-338. doi: 10.1161/CIRCULATIONAHA.122.060788. Epub 2022 Jul 25. Circulation. 2022. PMID: 35877835 No abstract available.
References
-
- Caforio ALP. Myocarditis: endomyocardial biopsy and circulating anti-heart autoantibodies are key to diagnosis and personalized etiology-directed treatment. European Heart Journal. 2021;42:1618–1620. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
