

Oxytocin receptors are widely distributed in the prairie vole (Microtus ochrogaster) brain: Relation to social behavior, genetic polymorphisms, and the dopamine system
- PMID: 35763609
- PMCID: PMC9474670
- DOI: 10.1002/cne.25382
Oxytocin receptors are widely distributed in the prairie vole (Microtus ochrogaster) brain: Relation to social behavior, genetic polymorphisms, and the dopamine system
Abstract
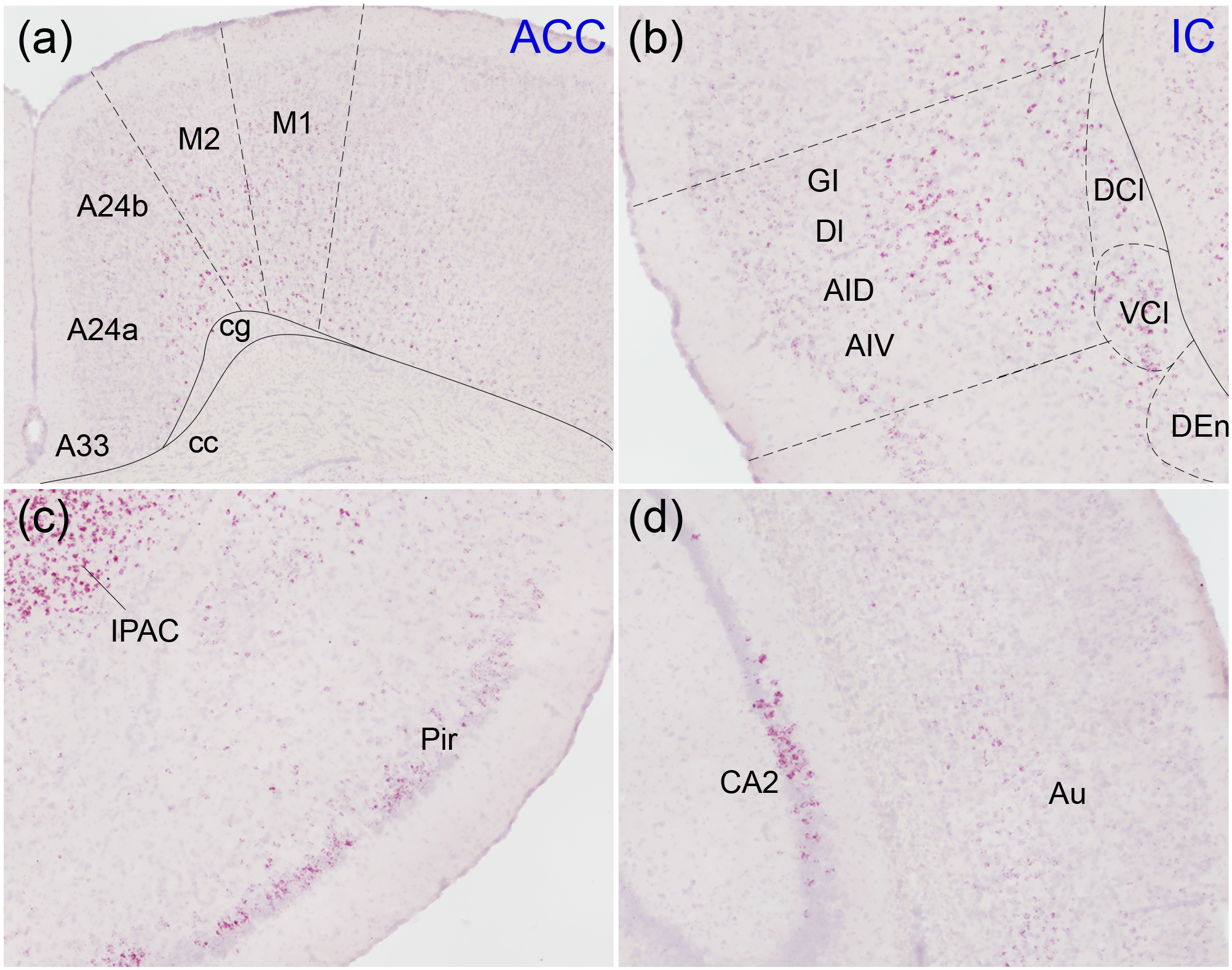
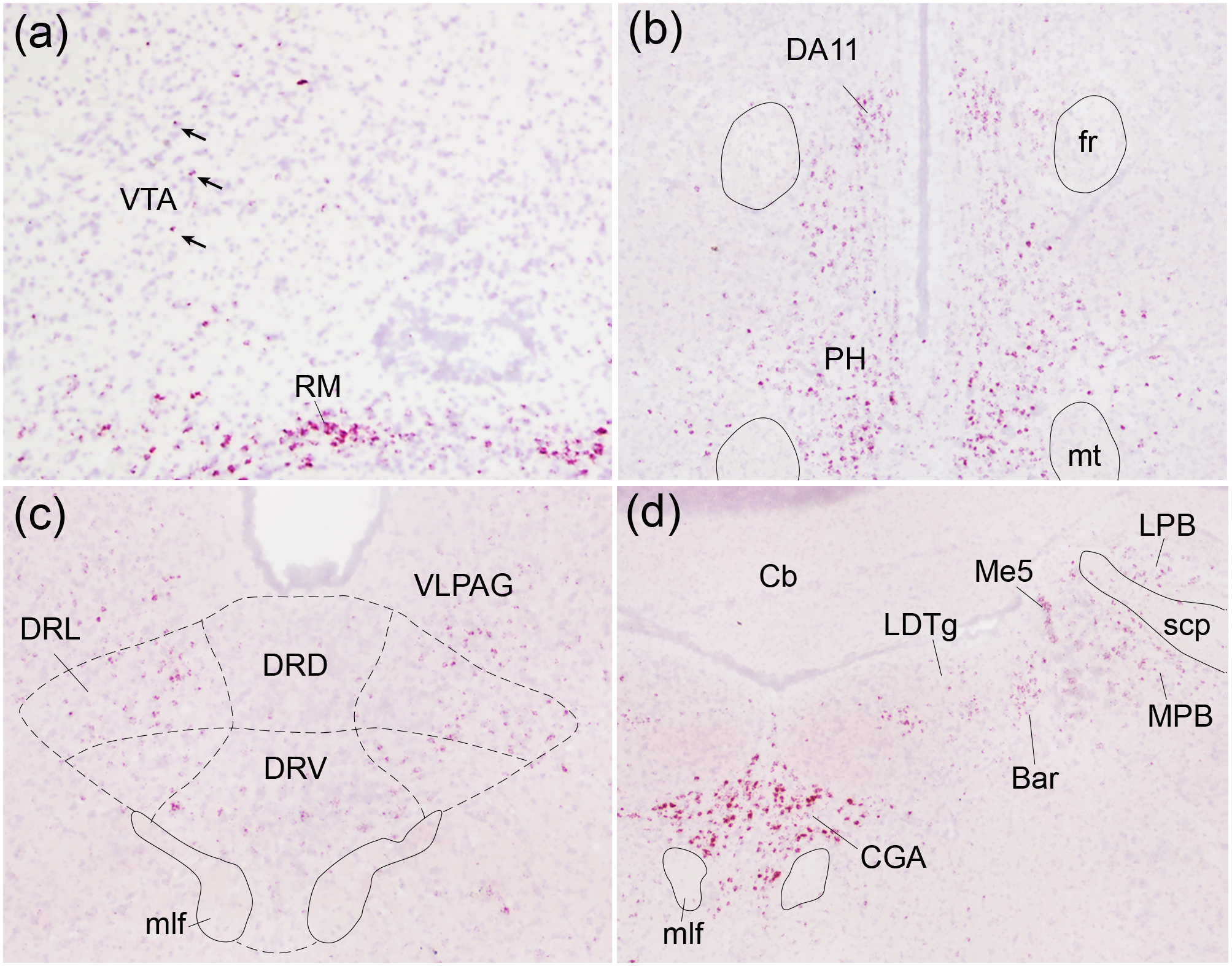
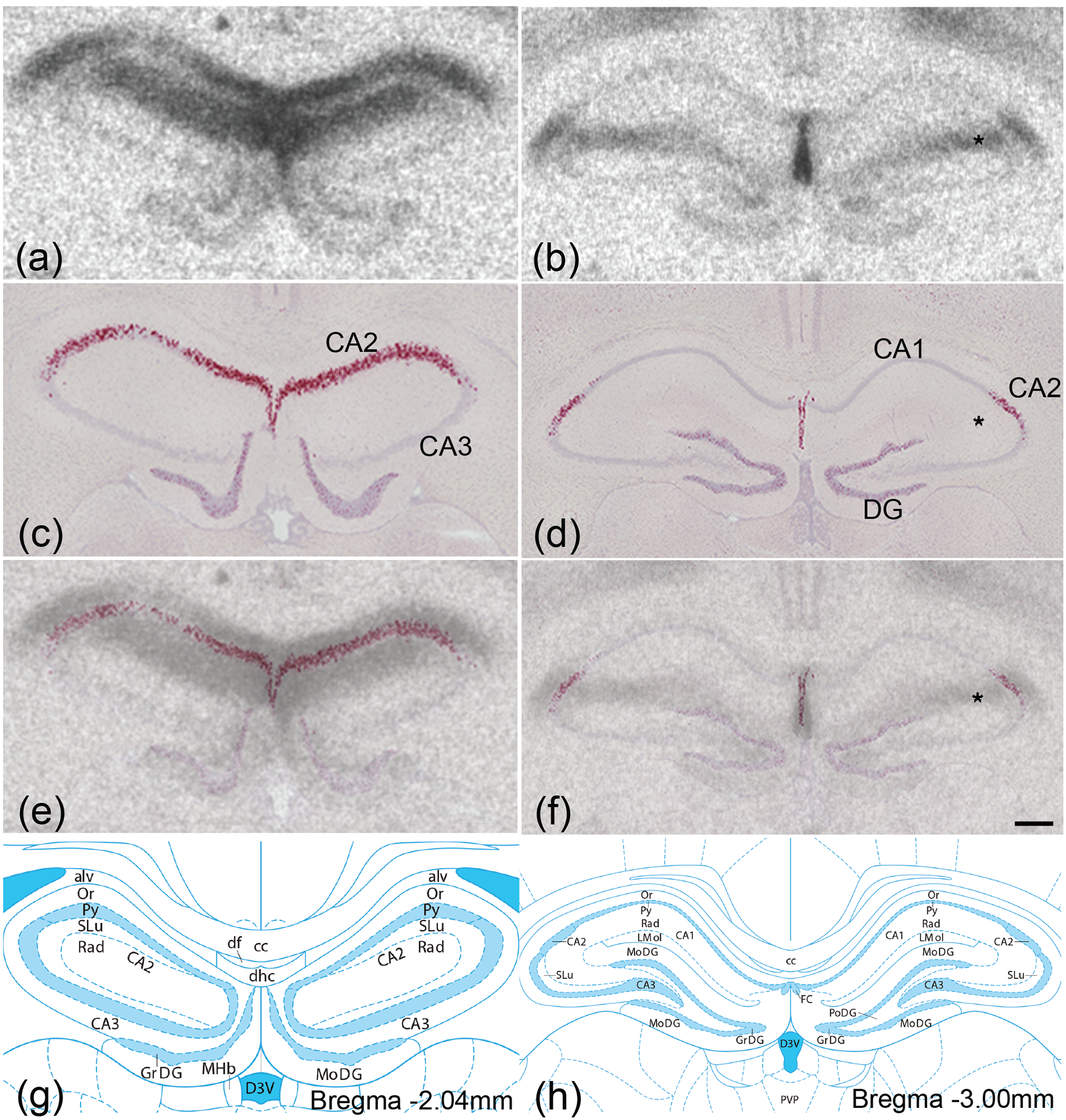
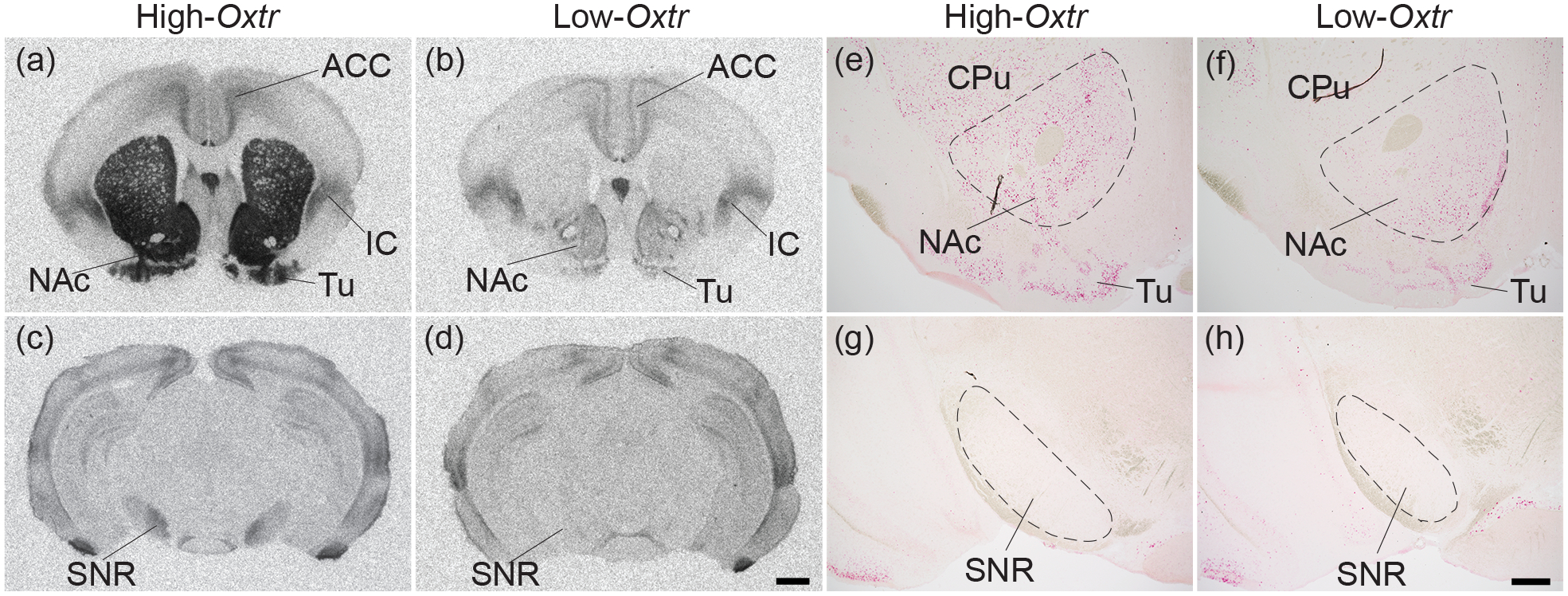
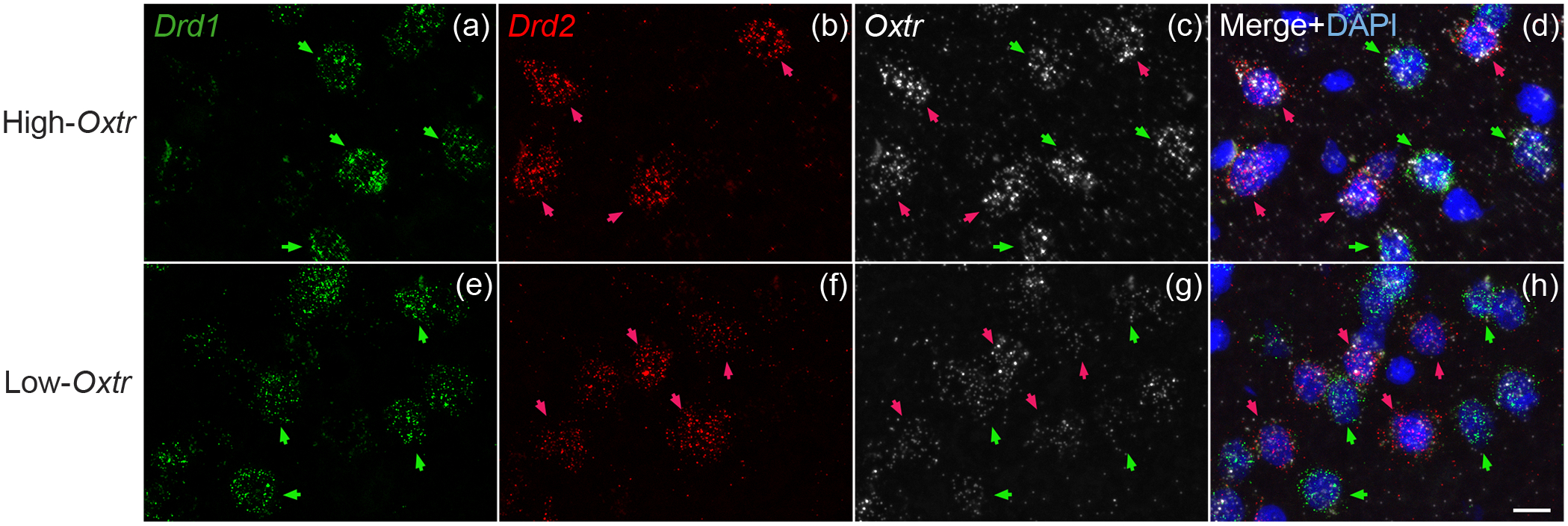
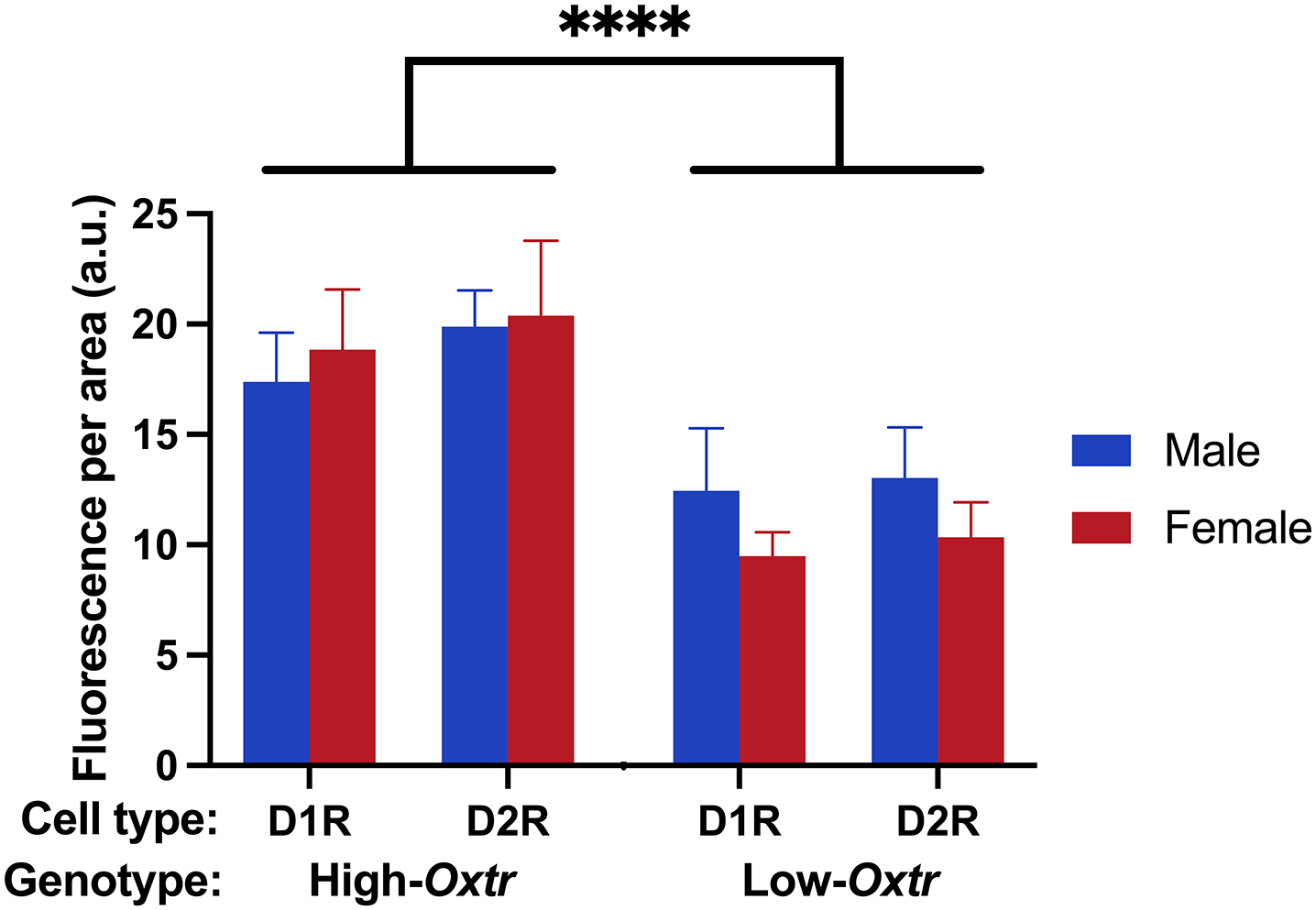
Oxytocin regulates social behavior via direct modulation of neurons, regulation of neural network activity, and interaction with other neurotransmitter systems. The behavioral effects of oxytocin signaling are determined by the species-specific distribution of brain oxytocin receptors. The socially monogamous prairie vole has been a useful model organism for elucidating the role of oxytocin in social behaviors, including pair bonding, response to social loss, and consoling. However, there has been no comprehensive mapping of oxytocin receptor-expressing cells throughout the prairie vole brain. Here, we employed a highly sensitive in situ hybridization, RNAscope, to construct an exhaustive, brain-wide map of oxytocin receptor mRNA-expressing cells. We found that oxytocin receptor mRNA expression was widespread and diffused throughout the brain, with specific areas displaying a particularly robust expression. Comparing receptor binding with mRNA revealed that regions of the hippocampus and substantia nigra contained oxytocin receptor protein but lacked mRNA, indicating that oxytocin receptors can be transported to distal neuronal processes, consistent with presynaptic oxytocin receptor functions. In the nucleus accumbens, a region involved in oxytocin-dependent social bonding, oxytocin receptor mRNA expression was detected in both the D1 and D2 dopamine receptor-expressing subtypes of cells. Furthermore, natural genetic polymorphisms robustly influenced oxytocin receptor expression in both D1 and D2 receptor cell types in the nucleus accumbens. Collectively, our findings further elucidate the extent to which oxytocin signaling is capable of influencing brain-wide neural activity, responses to social stimuli, and social behavior. KEY POINTS: Oxytocin receptor mRNA is diffusely expressed throughout the brain, with strong expression concentrated in certain areas involved in social behavior. Oxytocin receptor mRNA expression and protein localization are misaligned in some areas, indicating that the receptor protein may be transported to distal processes. In the nucleus accumbens, oxytocin receptors are expressed on cells expressing both D1 and D2 dopamine receptor subtypes, and the majority of variation in oxytocin receptor expression between animals is attributable to polymorphisms in the oxytocin receptor gene.
Keywords: Oxtr single nucleotide polymorphisms; autoradiography; dopamine receptor; in situ hybridization; nucleus accumbens; oxytocin receptors mRNA; phenotypic plasticity.
© 2022 Wiley Periodicals LLC.
Conflict of interest statement
Figures
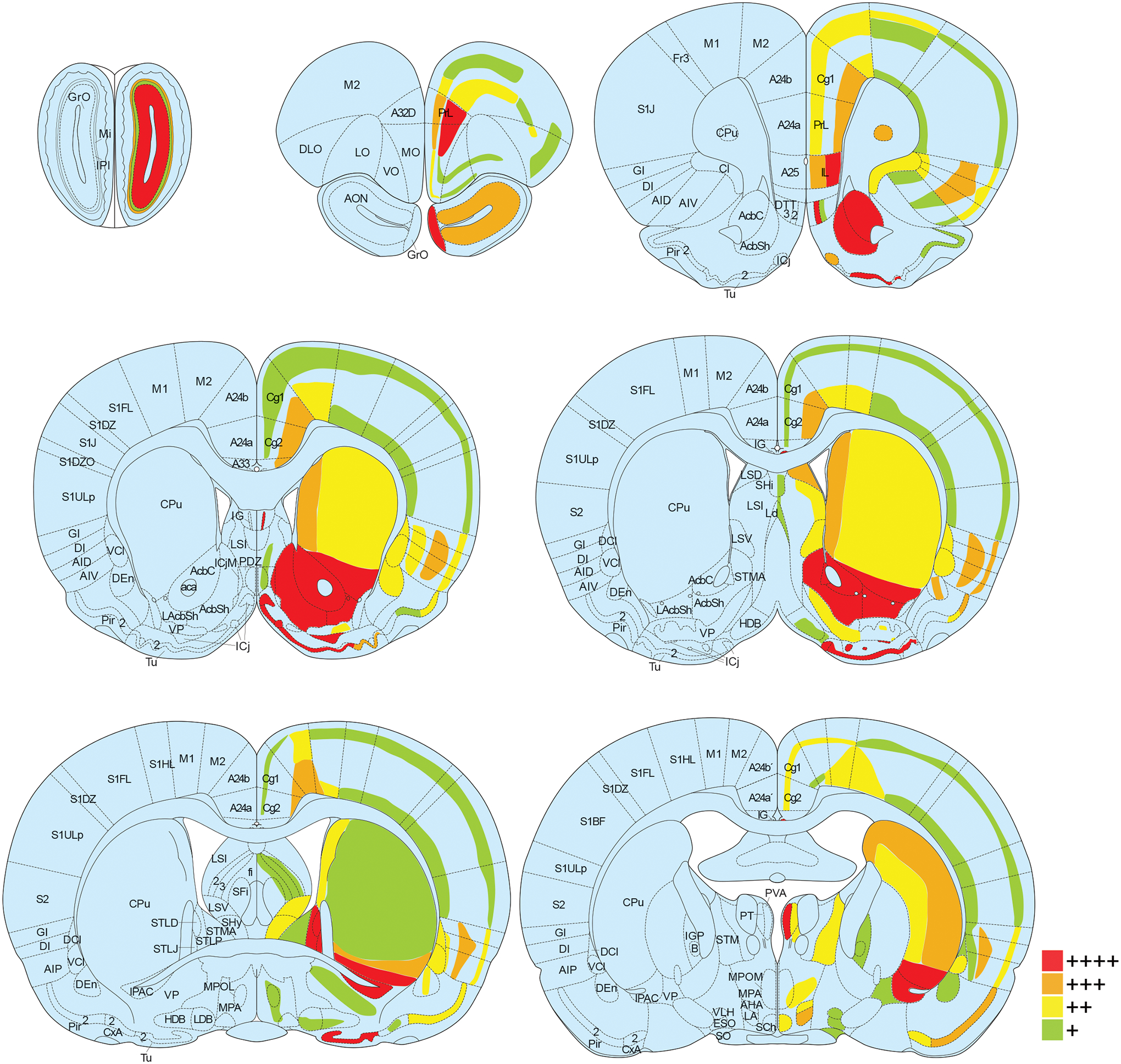
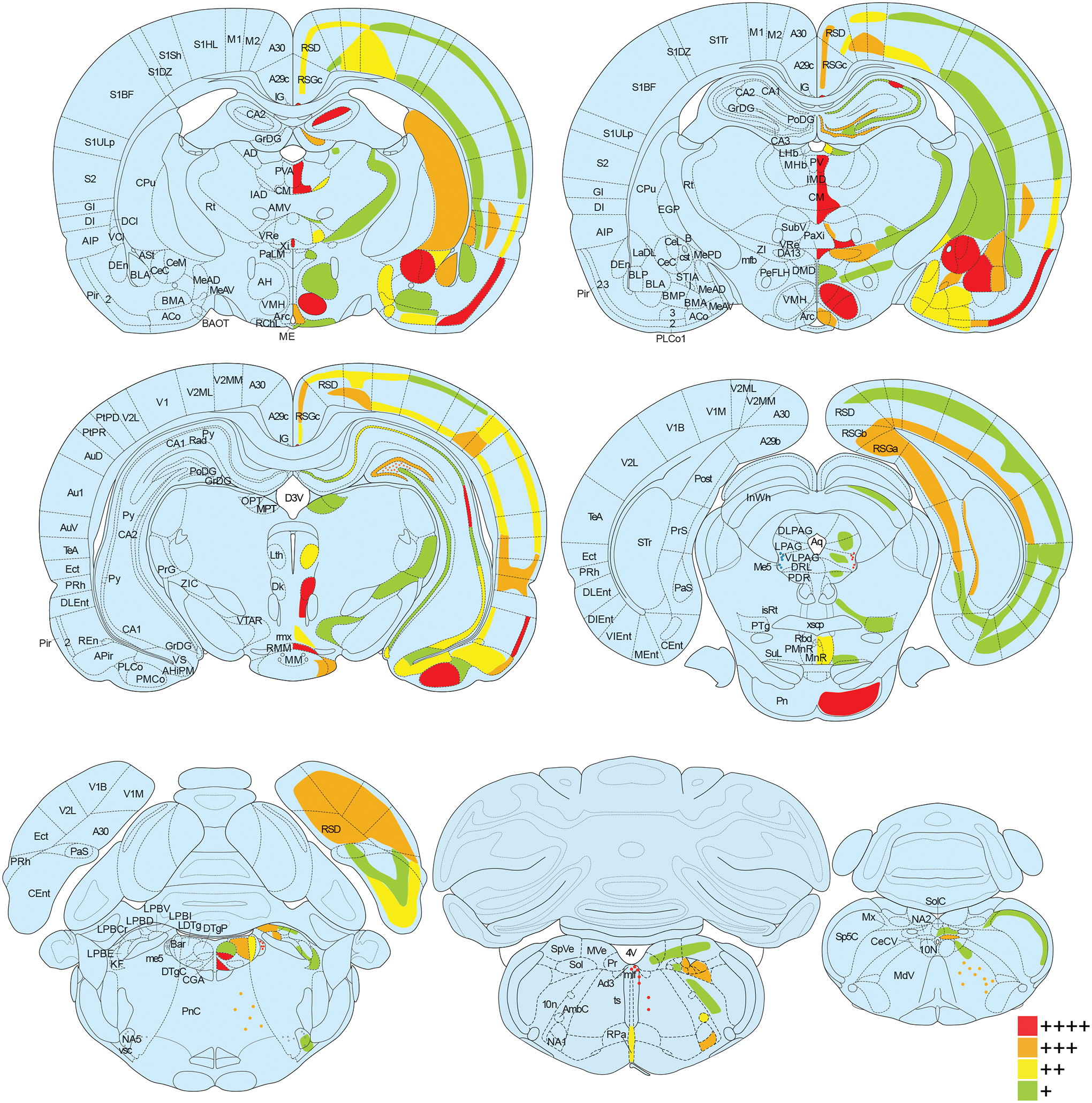
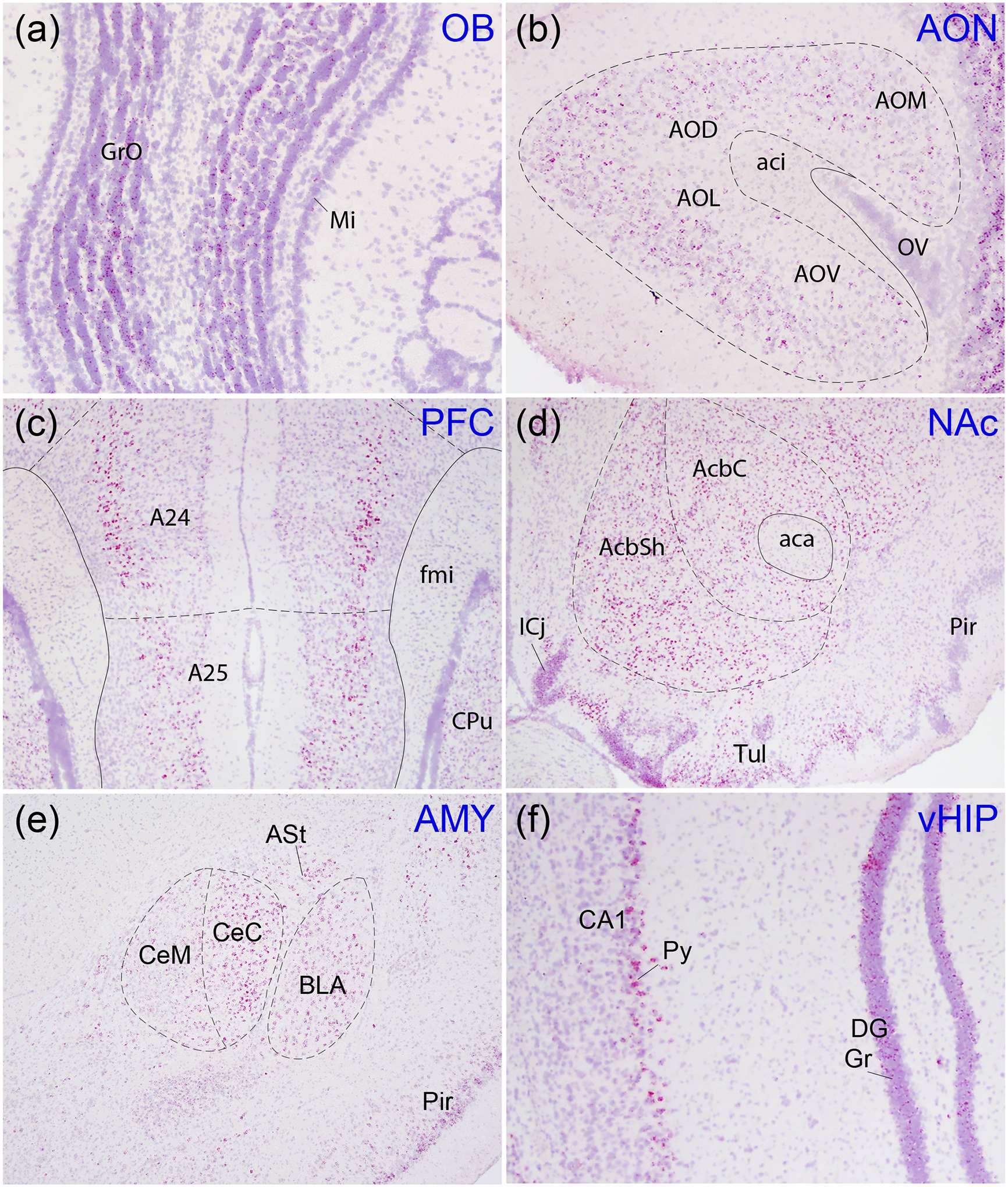
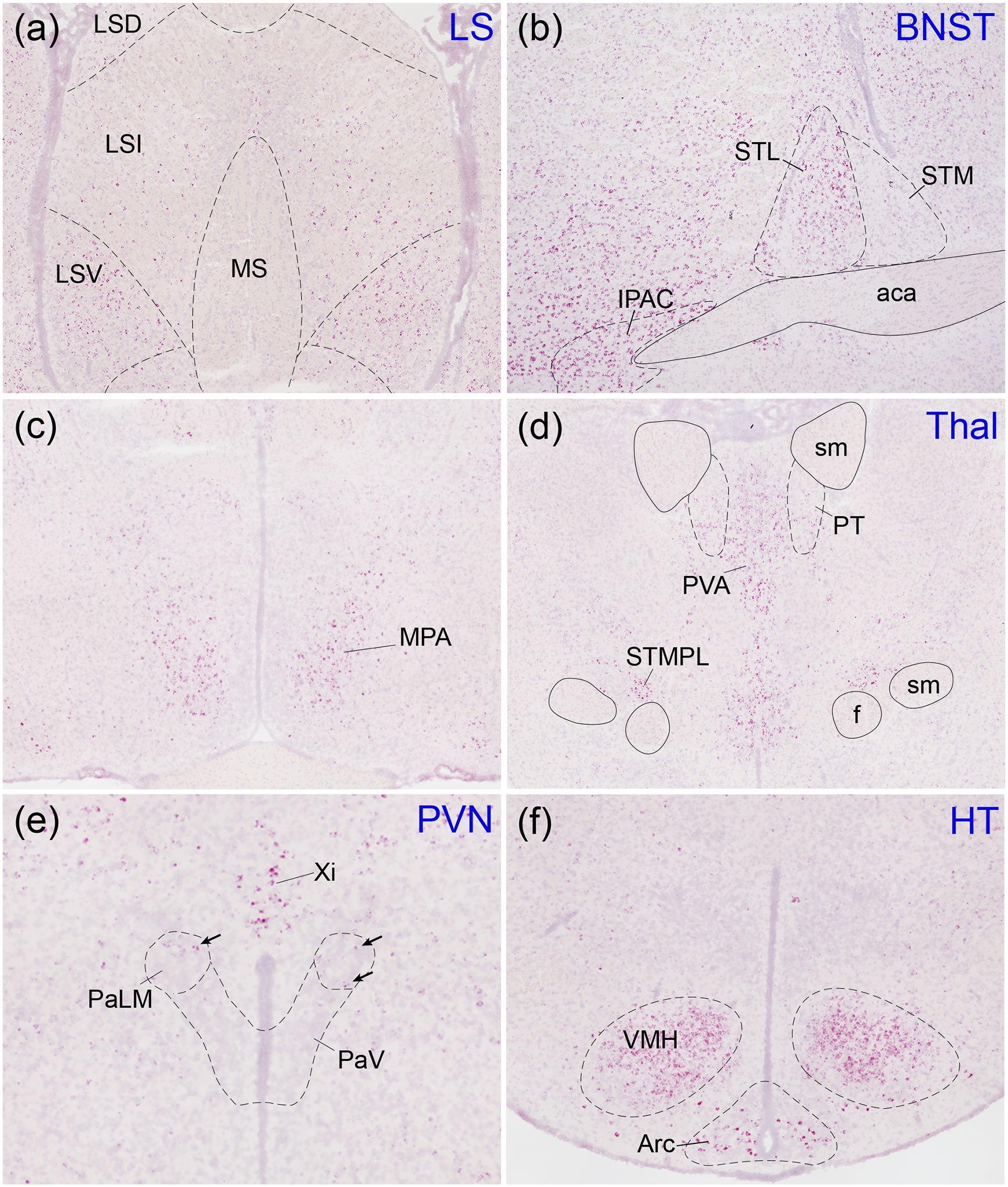
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
