

Differential expression patterns of phospholipase D isoforms 1 and 2 in the mammalian brain and retina
- PMID: 35764123
- PMCID: PMC9305353
- DOI: 10.1016/j.jlr.2022.100247
Differential expression patterns of phospholipase D isoforms 1 and 2 in the mammalian brain and retina
Abstract
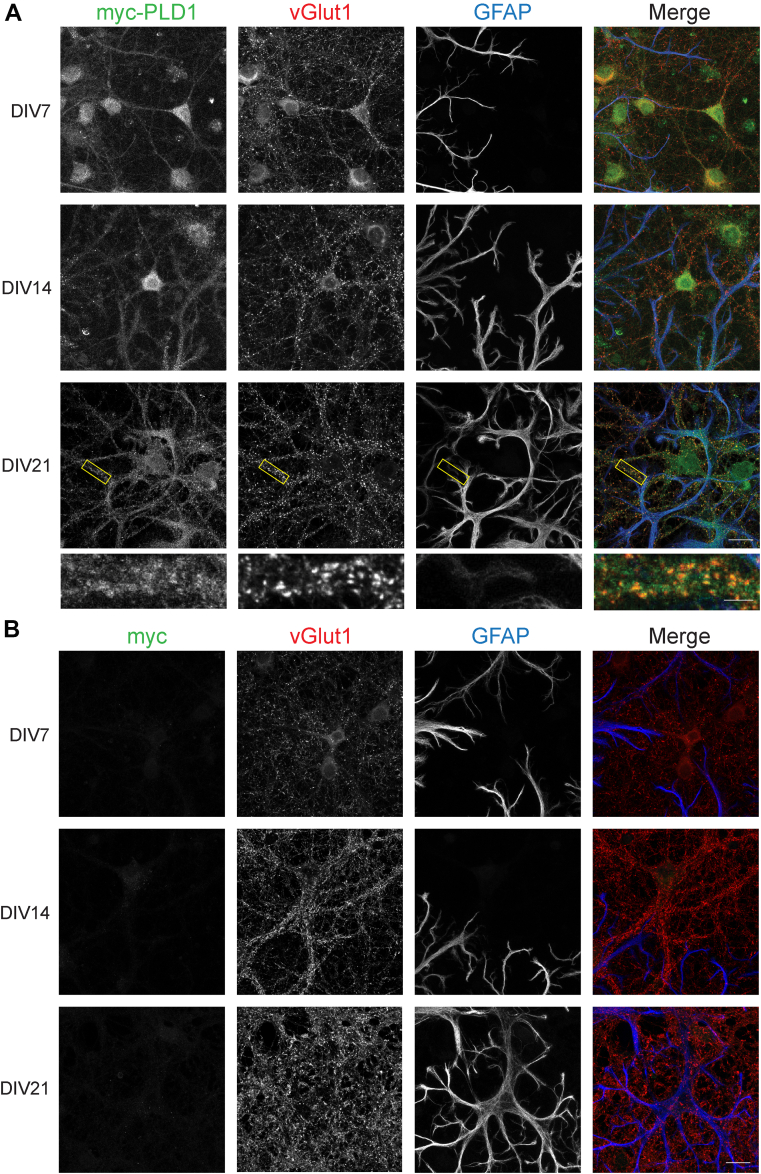
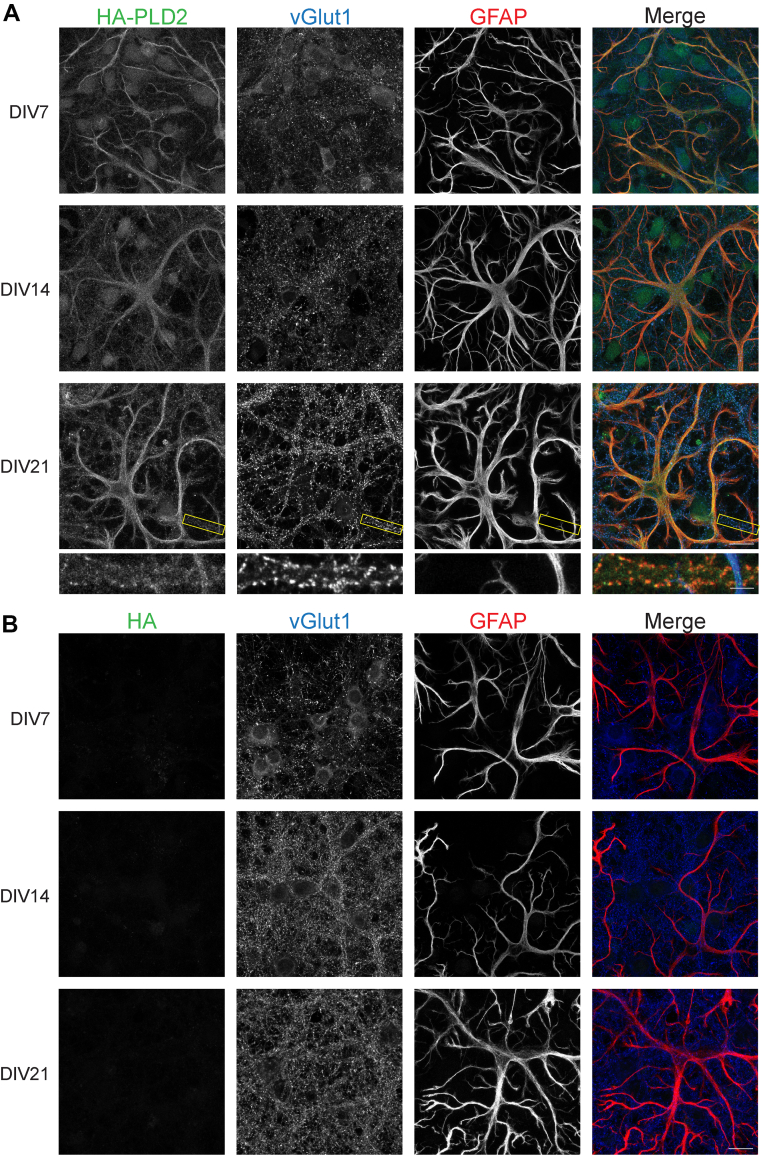
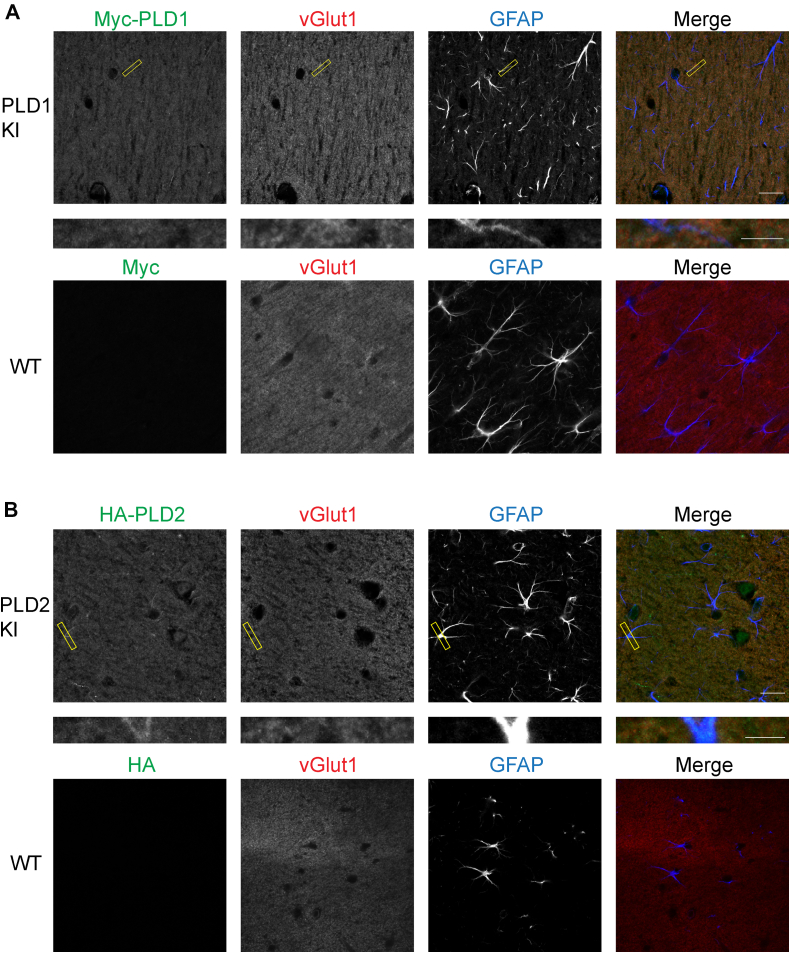
Phosphatidic acid is a key signaling molecule heavily implicated in exocytosis due to its protein-binding partners and propensity to induce negative membrane curvature. One phosphatidic acid-producing enzyme, phospholipase D (PLD), has also been implicated in neurotransmission. Unfortunately, due to the unreliability of reagents, there has been confusion in the literature regarding the expression of PLD isoforms in the mammalian brain which has hampered our understanding of their functional roles in neurons. To address this, we generated epitope-tagged PLD1 and PLD2 knockin mice using CRISPR/Cas9. Using these mice, we show that PLD1 and PLD2 are both localized at synapses by adulthood, with PLD2 expression being considerably higher in glial cells and PLD1 expression predominating in neurons. Interestingly, we observed that only PLD1 is expressed in the mouse retina, where it is found in the synaptic plexiform layers. These data provide critical information regarding the localization and potential role of PLDs in the central nervous system.
Keywords: CRISPR/Cas9; brain lipids; exocytosis; eye/retina; glia; knock-in mice; neurotransmission; phospholipases/D; phospholipids/phosphatidic acid; synaptic plexiform layer.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures






References
-
- Zeniou-Meyer M., Zabari N., Ashery U., Chasserot-Golaz S., Haeberle A., Demais V., et al. Phospholipase D1 production of phosphatidic acid at the plasma membrane promotes exocytosis of large dense core granules at a late stage. J. Biol. Chem. 2007;282:21746–21757. - PubMed
-
- Tanguy E., Coste de Bagneaux P., Kassas N., Ammar M., Wang Q., Haeberle A., et al. Mono- and poly-unsaturated phosphatidic acid regulate distinct steps of regulated exocytosis in neuroendocrine cells. Cell Rep. 2020;32:108026. - PubMed
-
- English D. Phosphatidic acid: a lipid messenger involved in intracellular and extracellular signalling. Cell Signal. 1996;8:341–347. - PubMed
-
- Wang X., Devaiah S., Zhang W., Welti R. Signaling functions of phosphatidic acid. Prog. Lipid Res. 2006;45:250–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
