

Reassessment of Neuronal Tau Distribution in Adult Human Brain and Implications for Tau Pathobiology
- PMID: 35765058
- PMCID: PMC9237980
- DOI: 10.1186/s40478-022-01394-9
Reassessment of Neuronal Tau Distribution in Adult Human Brain and Implications for Tau Pathobiology
Abstract
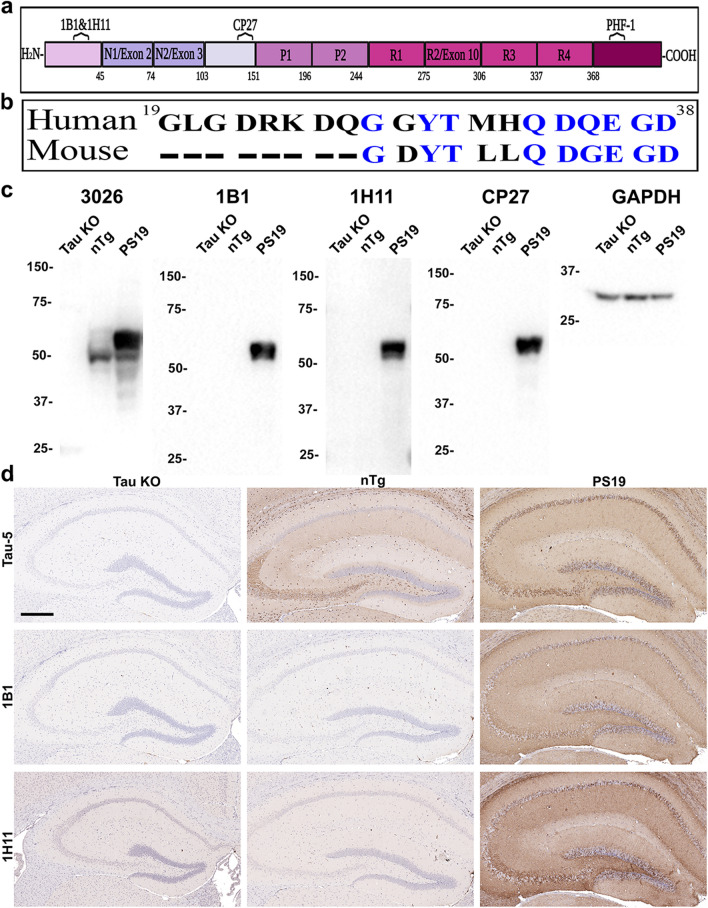
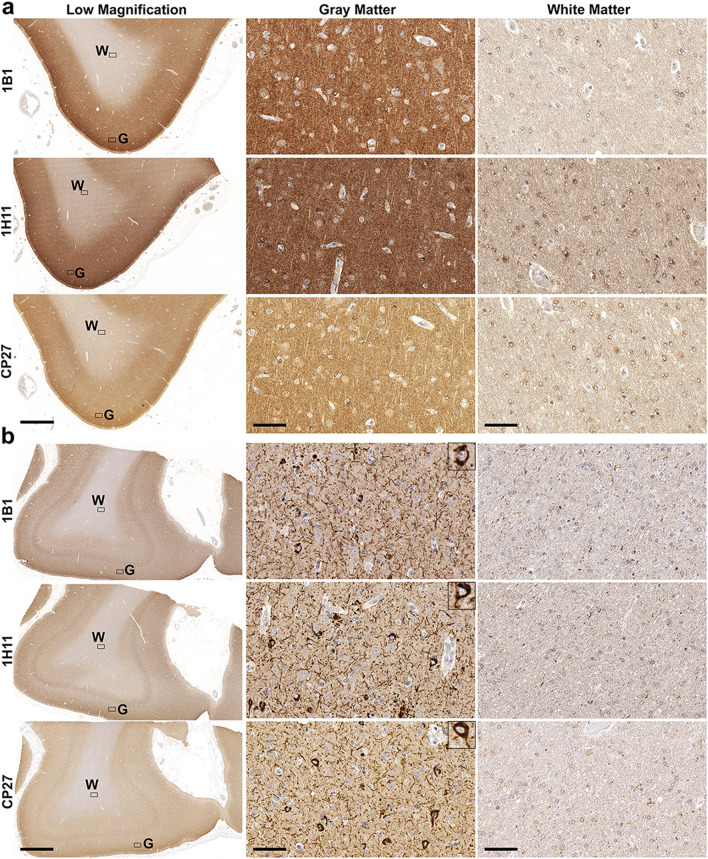
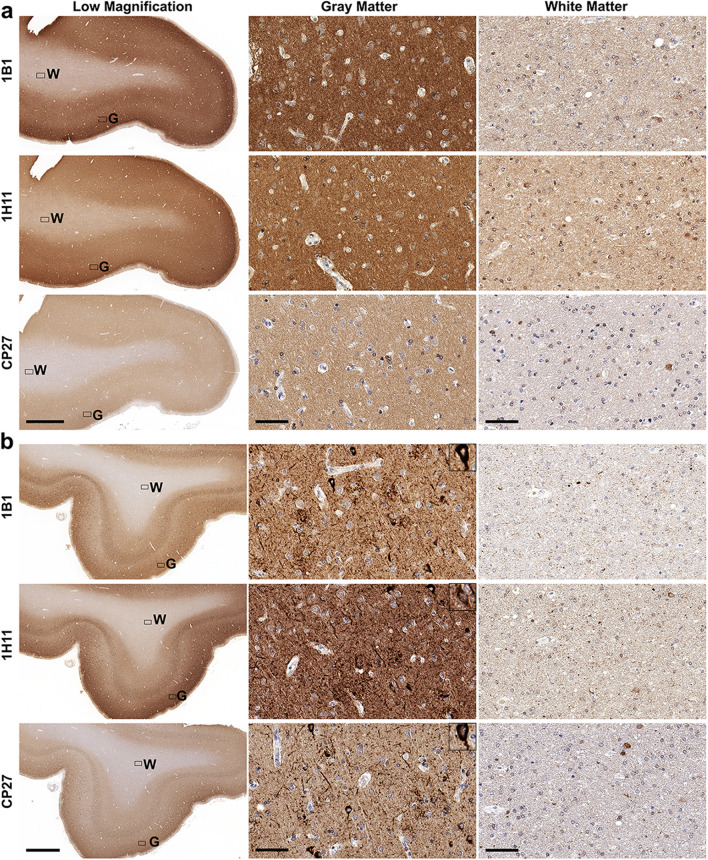
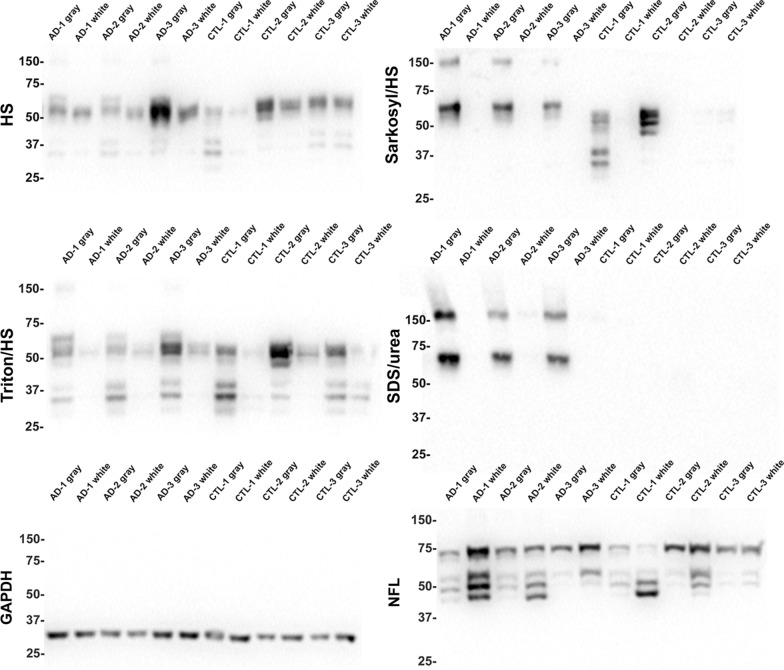
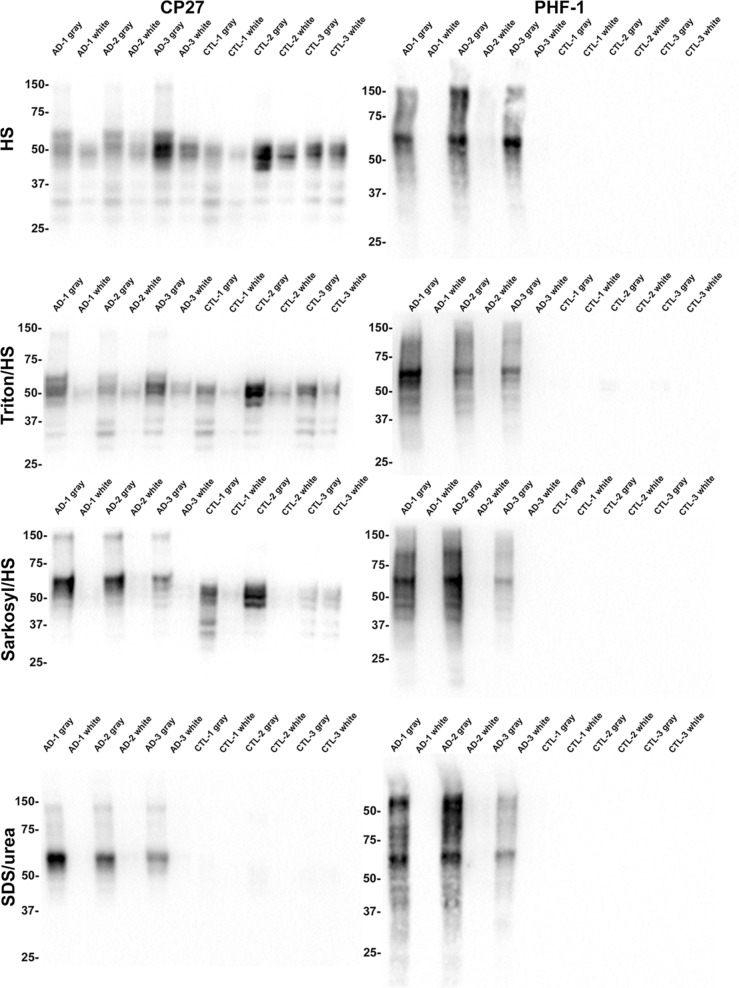
Tau is a predominantly neuronal, soluble and natively unfolded protein that can bind and stabilize microtubules in the central nervous system. Tau has been extensively studied over several decades, especially in the context of neurodegenerative diseases where it can aberrantly aggregate to form a spectrum of pathological inclusions. The presence of tau inclusions in the form of neurofibrillary tangles, neuropil threads and dystrophic neurites within senile plaques are essential and defining features of Alzheimer's disease. The current dogma favors the notion that tau is predominantly an axonal protein, and that in Alzheimer's disease there is a redistribution of tau towards the neuronal soma that is associated with the formation of pathological inclusions such as neurofibrillary tangles and neuropil threads. Using novel as well as previously established highly specific tau antibodies, we demonstrate that contrary to this overwhelmingly accepted fact, as asserted in numerous articles and reviews, in adult human brain, tau is more abundant in cortical gray matter that is enriched in neuronal soma and dendrites compared to white matter that is predominantly rich in neuronal axons. Additionally, in Alzheimer's disease tau pathology is significantly more abundant in the brain cortical gray matter of affected brain regions compared to the adjacent white matter regions. These findings have important implications for the biological function of tau as well as the mechanisms involved in the progressive spread of tau associated with the insidious nature of Alzheimer's disease.
Keywords: Alzheimer’s disease; Brain; Distribution; Human; Neuronal; Tau; Tauopathy.
© 2022. The Author(s).
Conflict of interest statement
We declare no conflict of interest in this manuscript.
Figures
References
-
- Aragão Gomes L, Uytterhoeven V, Lopez-Sanmartin D, Tomé SO, Tousseyn T, Vandenberghe R, Vandenbulcke M, von Arnim CAF, Verstreken P, Thal DR. Maturation of neuronal AD-tau pathology involves site-specific phosphorylation of cytoplasmic and synaptic tau preceding conformational change and fibril formation. Acta Neuropathol. 2021;141:173–192. doi: 10.1007/S00401-020-02251-6. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
