

Structural Characterization of L-Galactose Dehydrogenase: An Essential Enzyme for Vitamin C Biosynthesis
- PMID: 35765894
- PMCID: PMC9381564
- DOI: 10.1093/pcp/pcac090
Structural Characterization of L-Galactose Dehydrogenase: An Essential Enzyme for Vitamin C Biosynthesis
Abstract
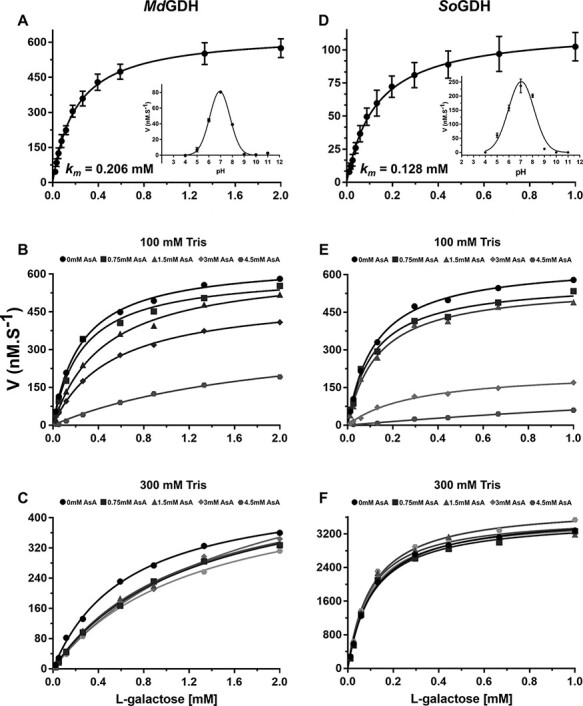
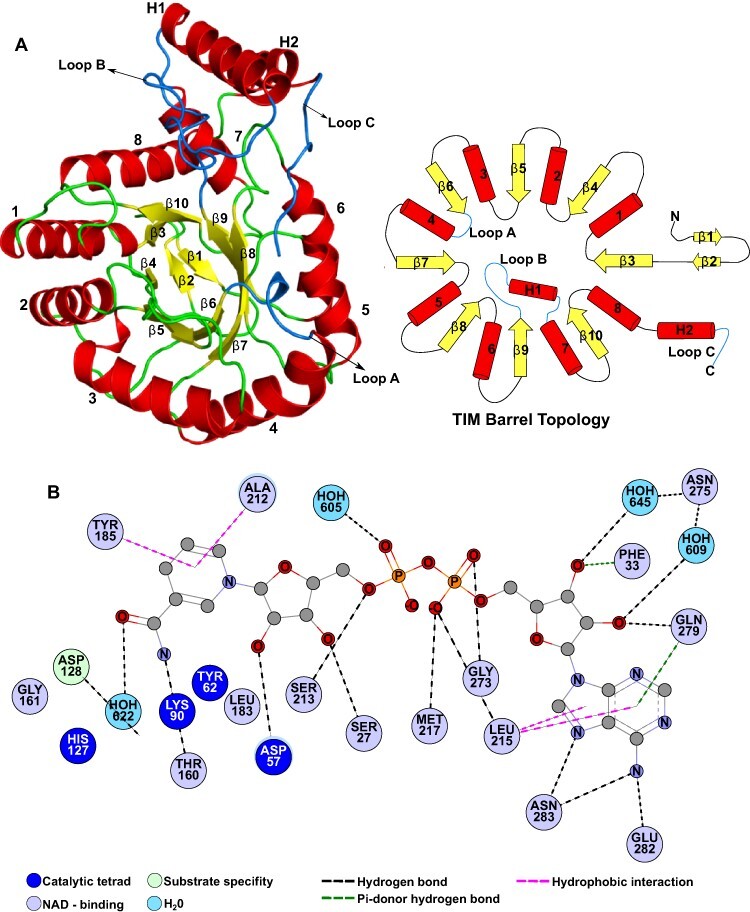
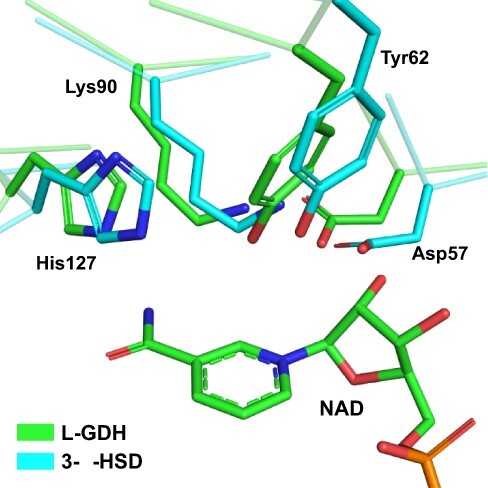
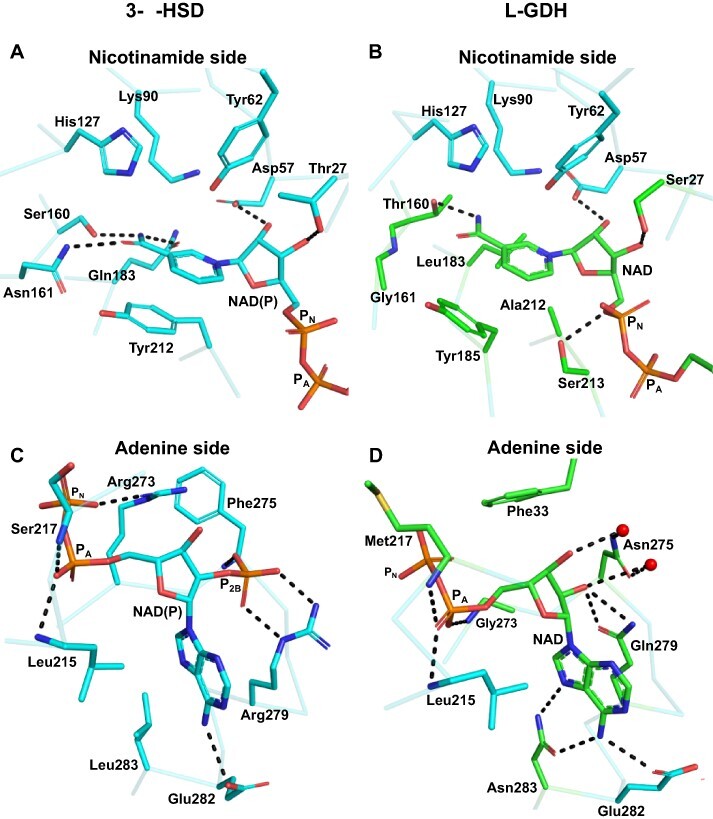
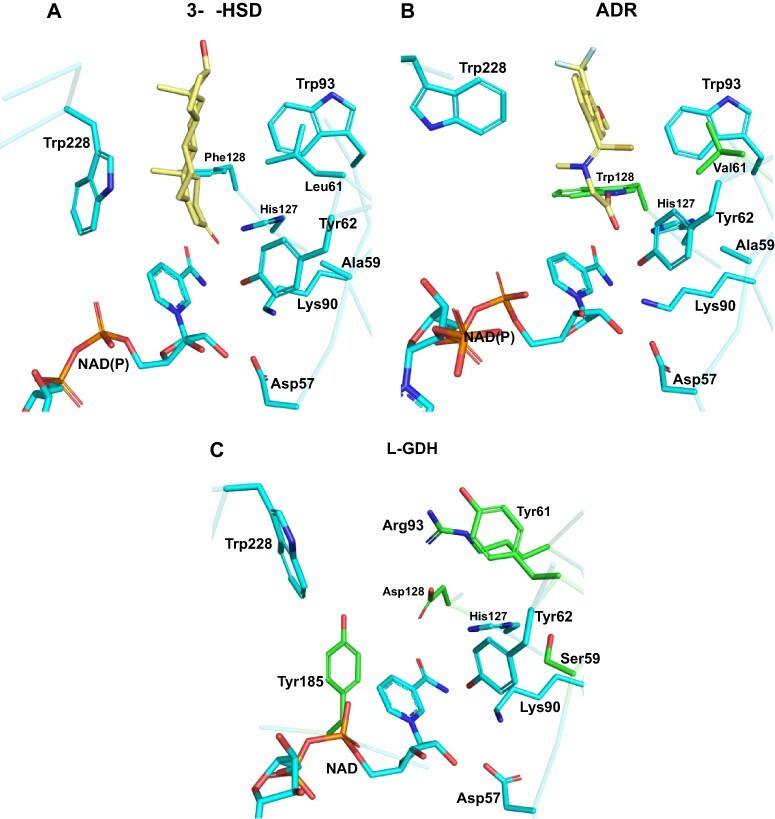
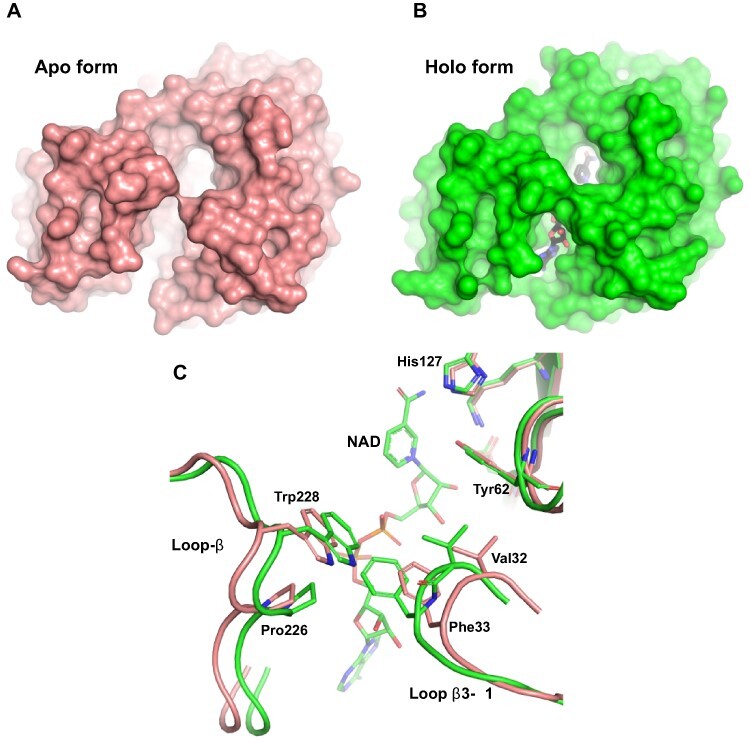
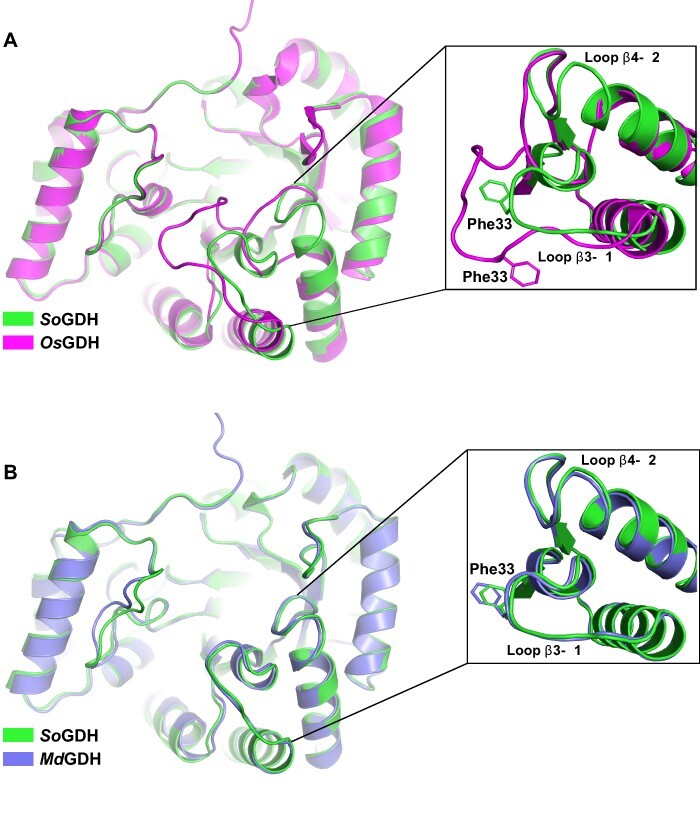
In plants, it is well-known that ascorbic acid (vitamin C) can be synthesized via multiple metabolic pathways but there is still much to be learned concerning their integration and control mechanisms. Furthermore, the structural biology of the component enzymes has been poorly exploited. Here we describe the first crystal structure for an L-galactose dehydrogenase [Spinacia oleracea GDH (SoGDH) from spinach], from the D-mannose/L-galactose (Smirnoff-Wheeler) pathway which converts L-galactose into L-galactono-1,4-lactone. The kinetic parameters for the enzyme are similar to those from its homolog from camu camu, a super-accumulator of vitamin C found in the Peruvian Amazon. Both enzymes are monomers in solution and have a pH optimum of 7, and their activity is largely unaffected by high concentrations of ascorbic acid, suggesting the absence of a feedback mechanism acting via GDH. Previous reports may have been influenced by changes of the pH of the reaction medium as a function of ascorbic acid concentration. The structure of SoGDH is dominated by a (β/α)8 barrel closely related to aldehyde-keto reductases (AKRs). The structure bound to NAD+ shows that the lack of Arg279 justifies its preference for NAD+ over NADP+, as employed by many AKRs. This favors the oxidation reaction that ultimately leads to ascorbic acid accumulation. When compared with other AKRs, residue substitutions at the C-terminal end of the barrel (Tyr185, Tyr61, Ser59 and Asp128) can be identified to be likely determinants of substrate specificity. The present work contributes toward a more comprehensive understanding of structure-function relationships in the enzymes involved in vitamin C synthesis.
Keywords: Myrciaria dubia ‘camu-camu’; Crystal structure; Enzyme kinetics; L-galactose dehydrogenase; Spinach; Vitamin C biosynthesis.
© The Author(s) 2022. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.
Figures

References
-
- Bennett M.J., Albert R.H., Jez J.M., Ma H., Penning T.M. and Lewis M. (1997) Steroid recognition and regulation of hormone action: crystal structure of testosterone and NADP+ bound to 3α-hydroxysteroid/dihydrodiol dehydrogenase. Structure 5: 799–812. - PubMed
MeSH terms
Substances
Grants and funding
- 1152-2020-UNAP/Universidad Nacional de la Amazonia Peruana (UNAP)
- 188-2018-INIA-PNIA-PASANTIA/Programa Nacional de Innovación Agraria (PNIA)
- 069-2019-FONDECYT/Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica (CONCYTEC)
- 88887.505769/2020-00/Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
LinkOut - more resources
Full Text Sources
