

A surface pocket in the cytoplasmic domain of the herpes simplex virus fusogen gB controls membrane fusion
- PMID: 35767585
- PMCID: PMC9275723
- DOI: 10.1371/journal.ppat.1010435
A surface pocket in the cytoplasmic domain of the herpes simplex virus fusogen gB controls membrane fusion
Abstract
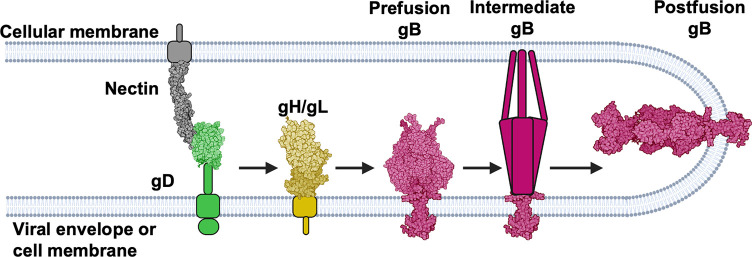
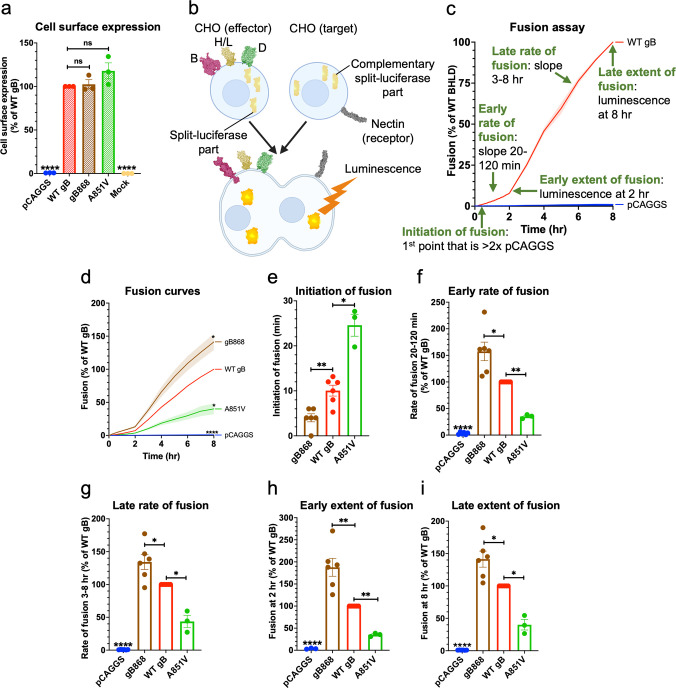
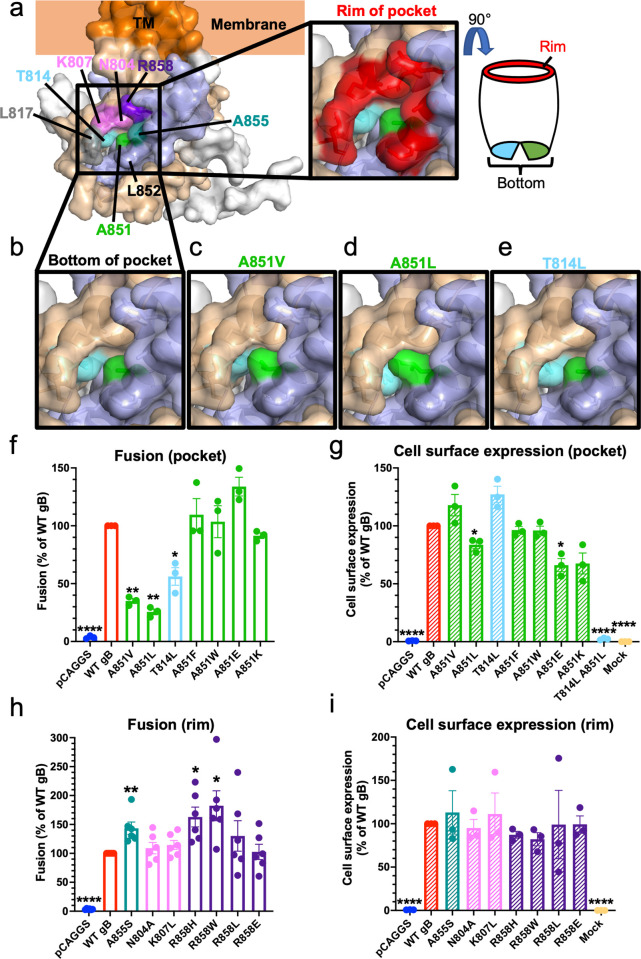
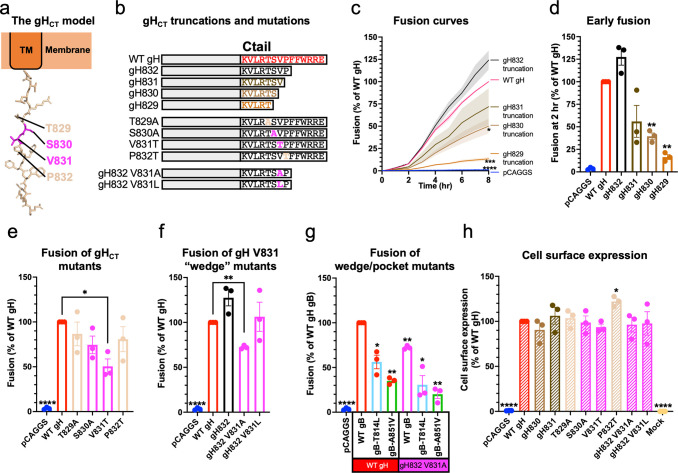
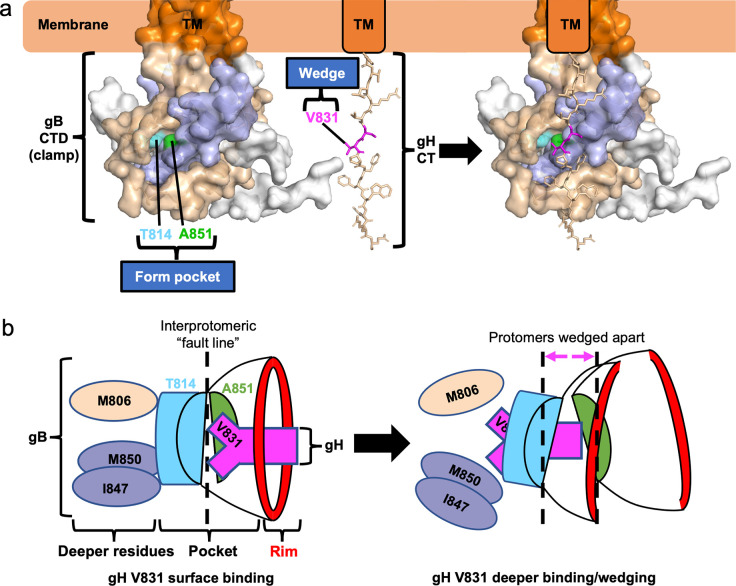
Membrane fusion during the entry of herpesviruses is carried out by the viral fusogen gB that is activated by its partner protein gH in some manner. The fusogenic activity of gB is controlled by its cytoplasmic (or intraviral) domain (gBCTD) and, according to the current model, the gBCTD is a trimeric, inhibitory clamp that restrains gB in the prefusion conformation. But how the gBCTD clamp is released by gH is unclear. Here, we identified two new regulatory elements within gB and gH from the prototypical herpes simplex virus 1: a surface pocket within the gBCTD and residue V831 within the gH cytoplasmic tail. Mutagenesis and structural modeling suggest that gH V831 interacts with the gB pocket. The gB pocket is located above the interface between adjacent protomers, and we hypothesize that insertion of the gH V831 wedge into the pocket serves to push the protomers apart, which releases the inhibitory clamp. In this manner, gH activates the fusogenic activity of gB. Both gB and gH are conserved across all herpesviruses, and this activation mechanism could be used by other gB homologs. Our proposed mechanism emphasizes a central role for the cytoplasmic regions in regulating the activity of a viral fusogen.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Cooper RS, Georgieva ER, Borbat PP, Freed JH, Heldwein EE. Structural basis for membrane anchoring and fusion regulation of the herpes simplex virus fusogen gB. Nat Struct Mol Biol. 2018;25(5):416–24. Epub 2018/05/08. doi: 10.1038/s41594-018-0060-6 ; PubMed Central PMCID: PMC5942590. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
