

Loss of Stathmin-2, a hallmark of TDP-43-associated ALS, causes motor neuropathy
- PMID: 35767949
- PMCID: PMC9327139
- DOI: 10.1016/j.celrep.2022.111001
Loss of Stathmin-2, a hallmark of TDP-43-associated ALS, causes motor neuropathy
Abstract
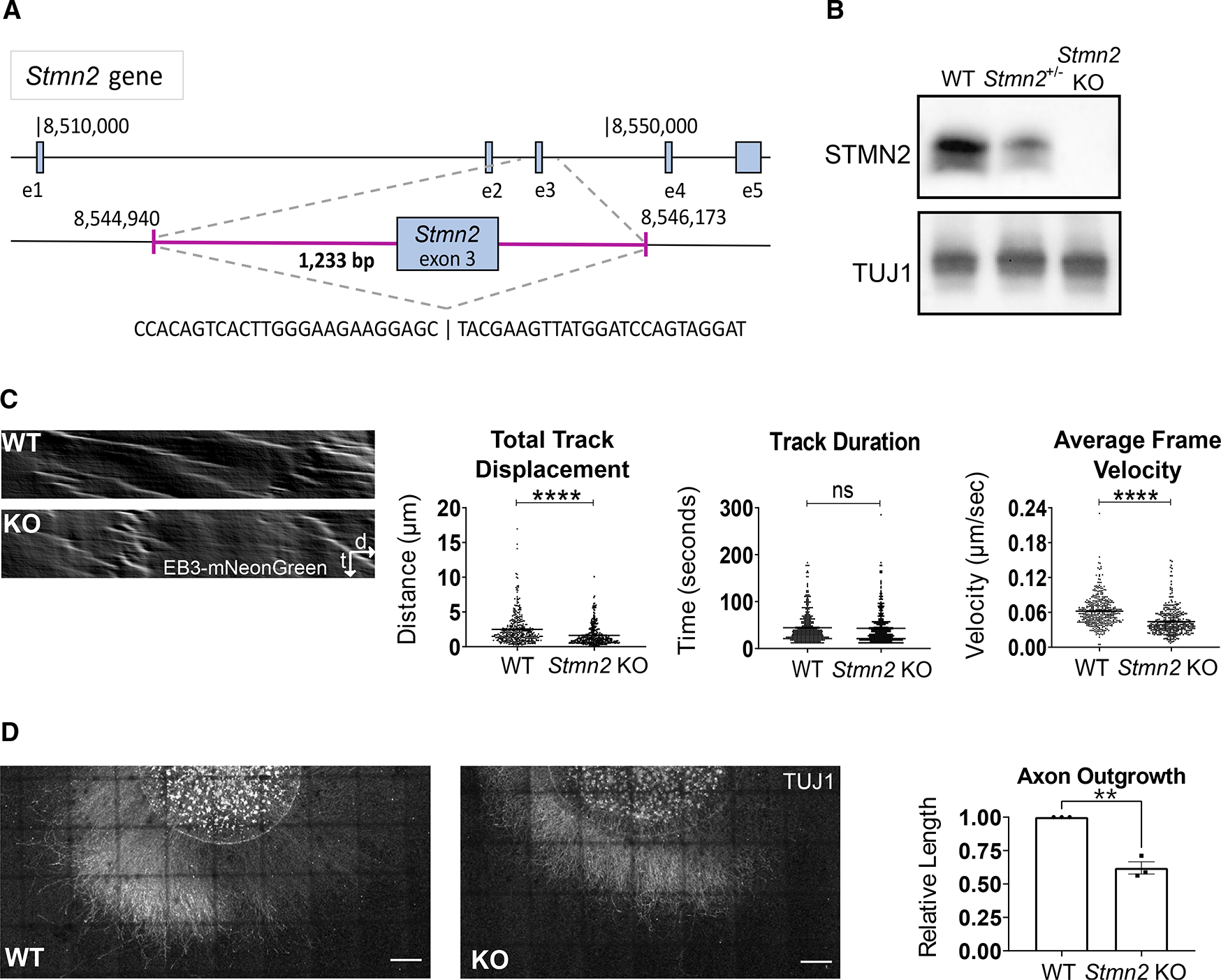
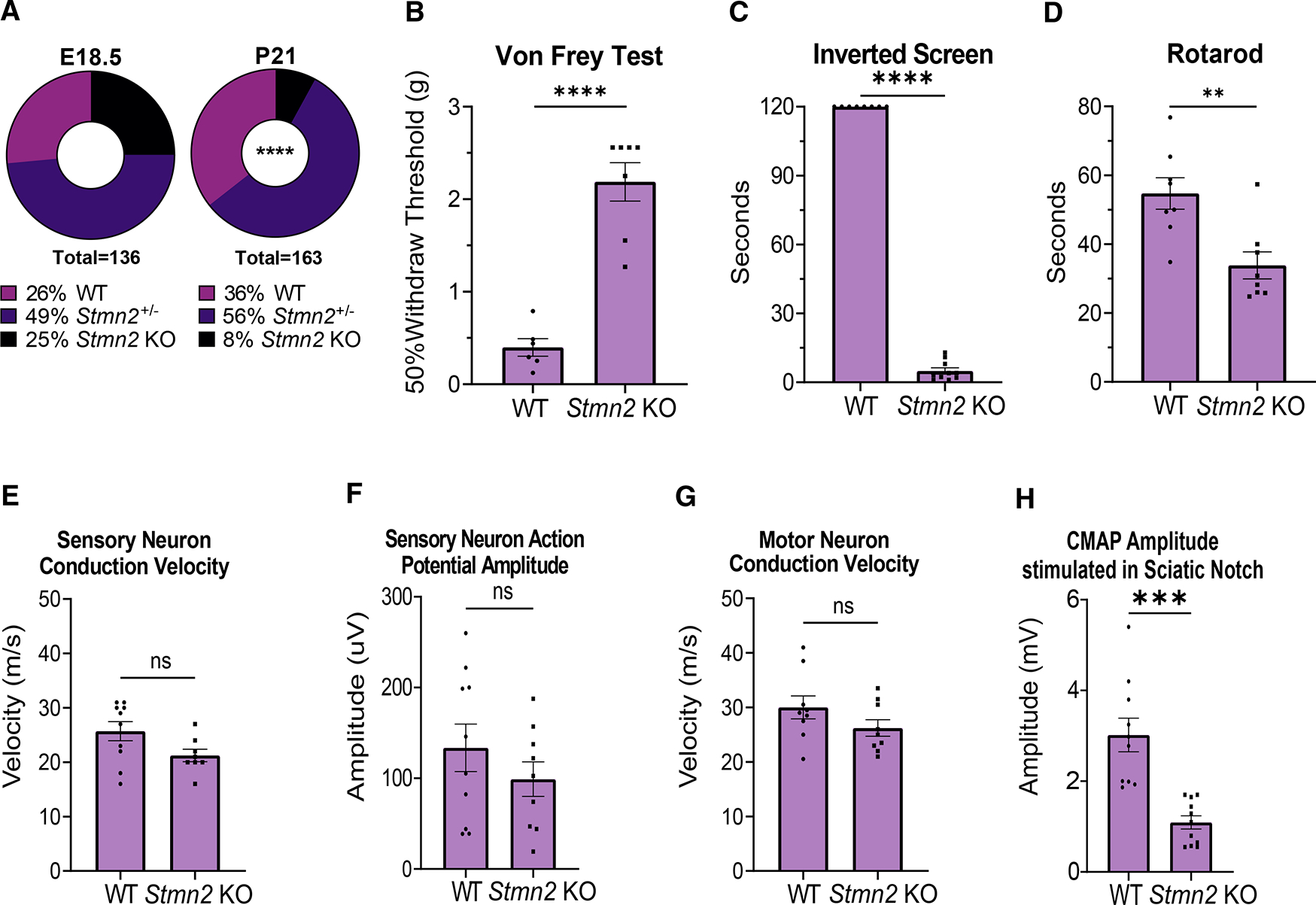
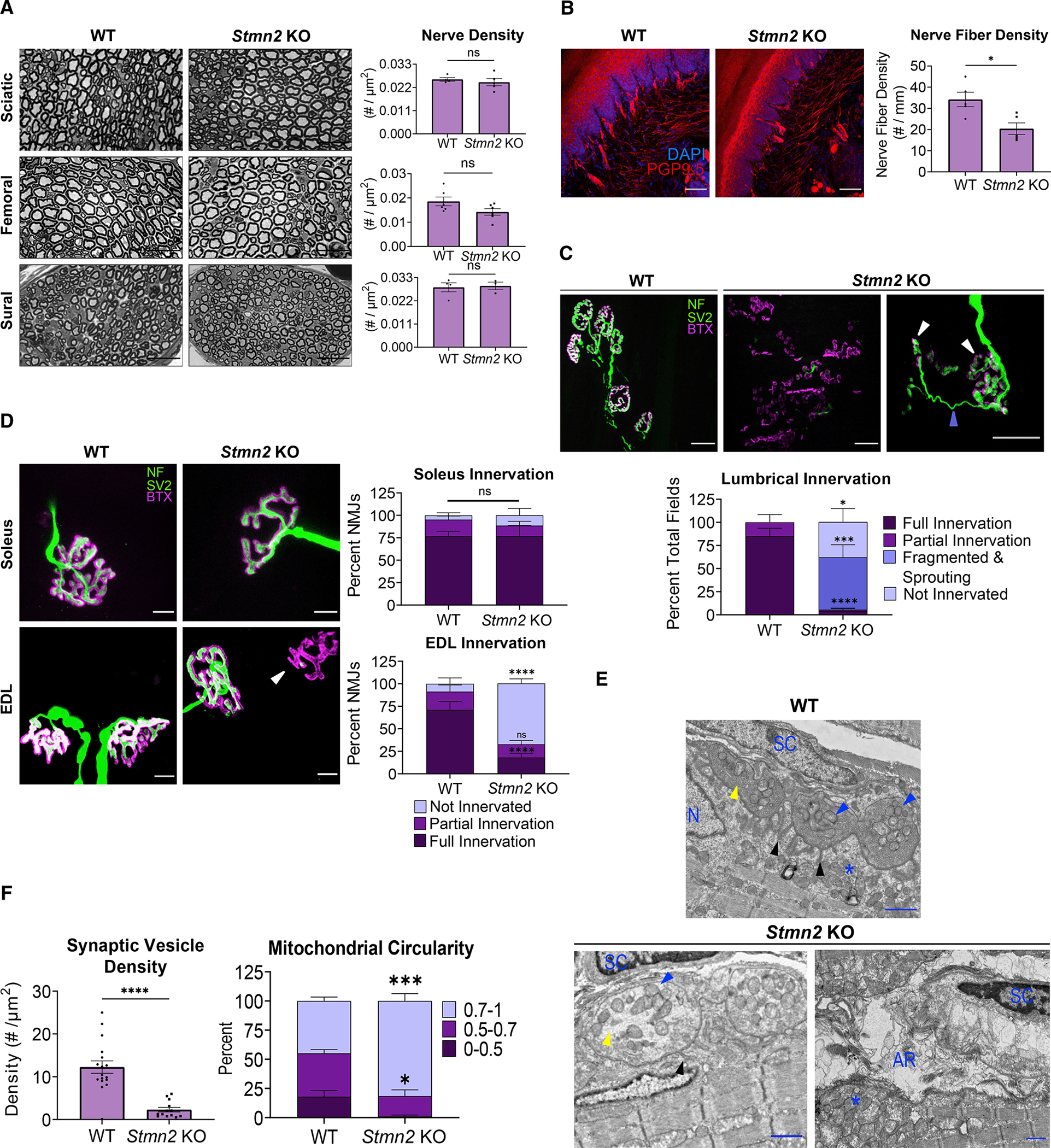
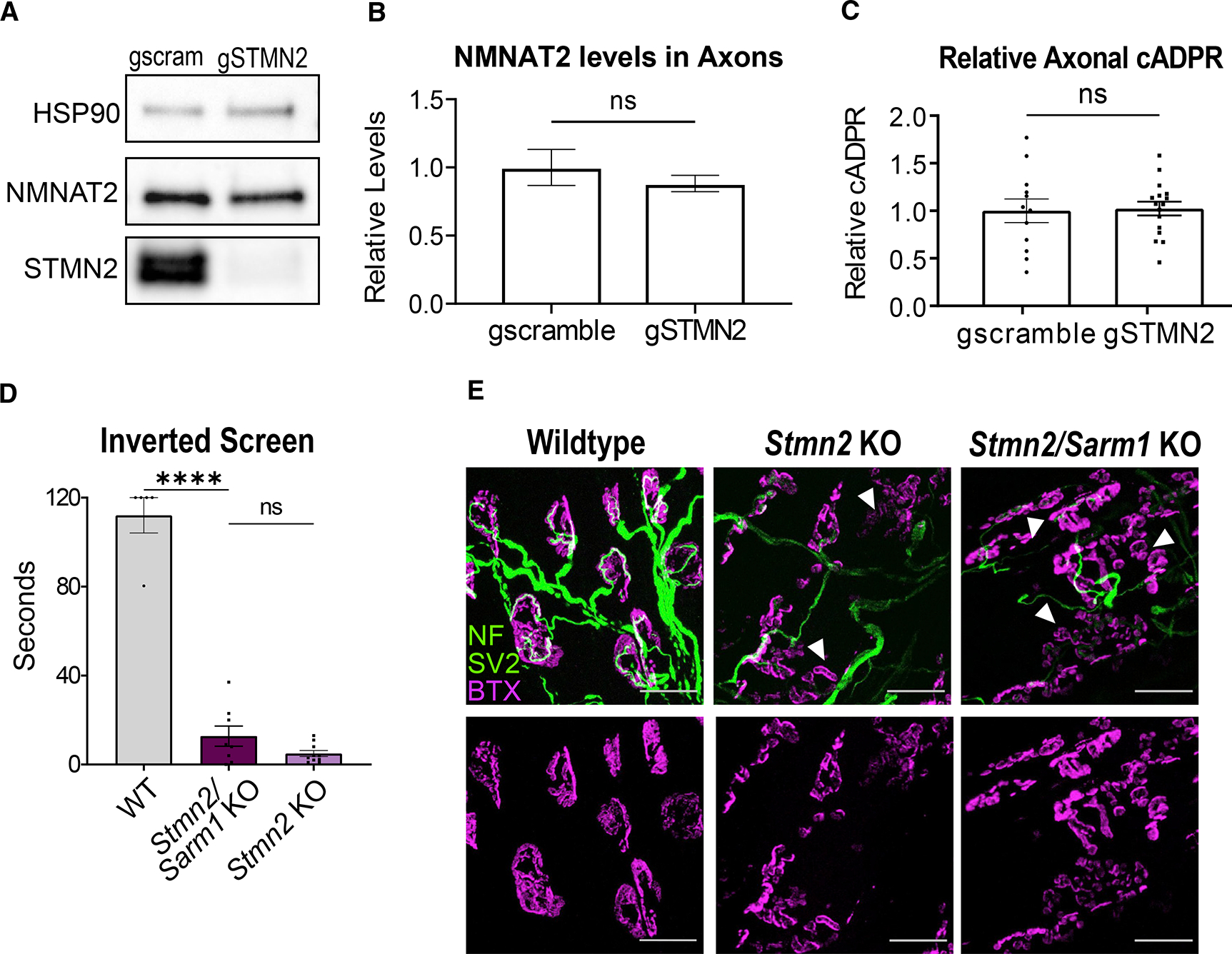
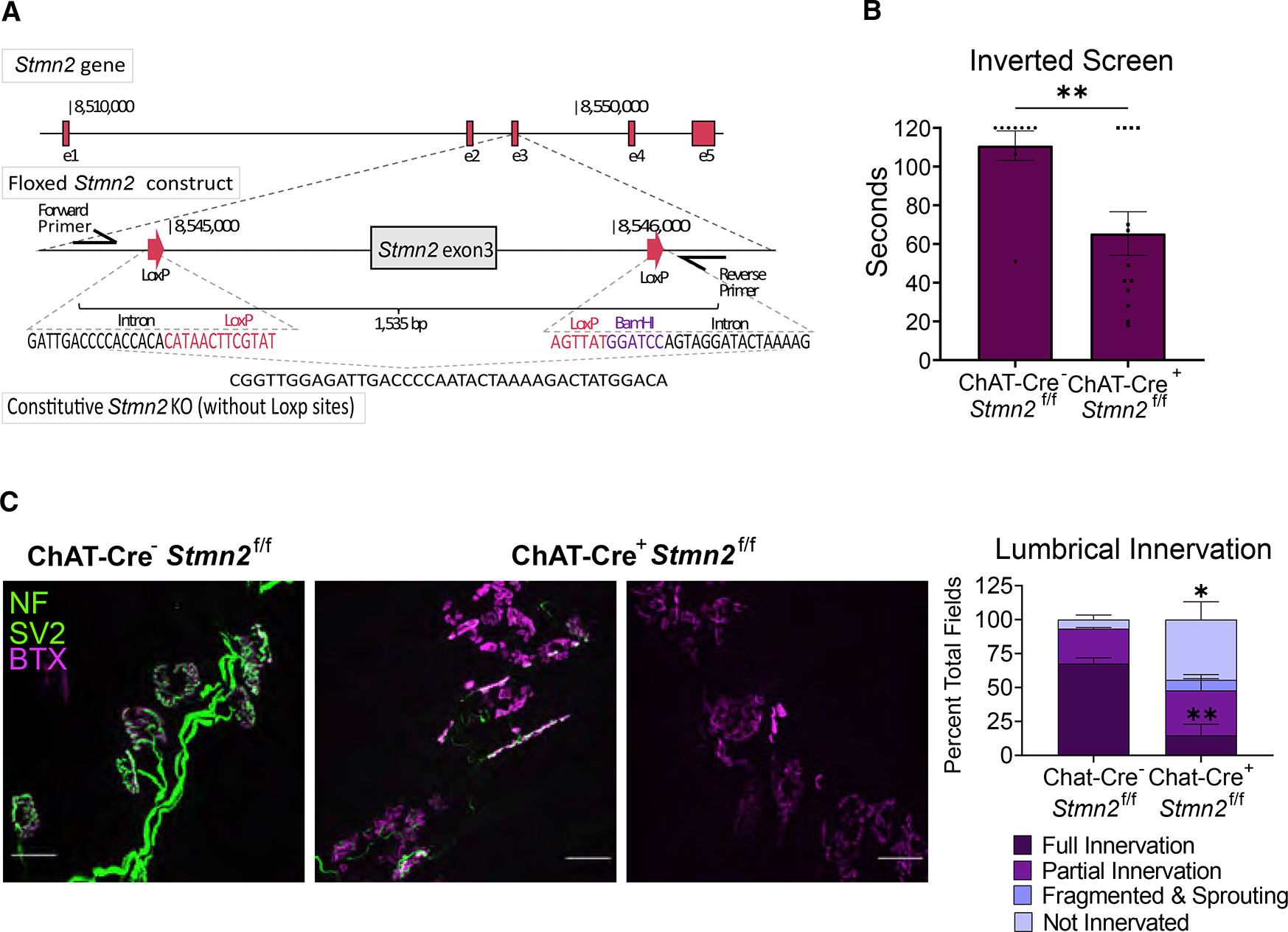
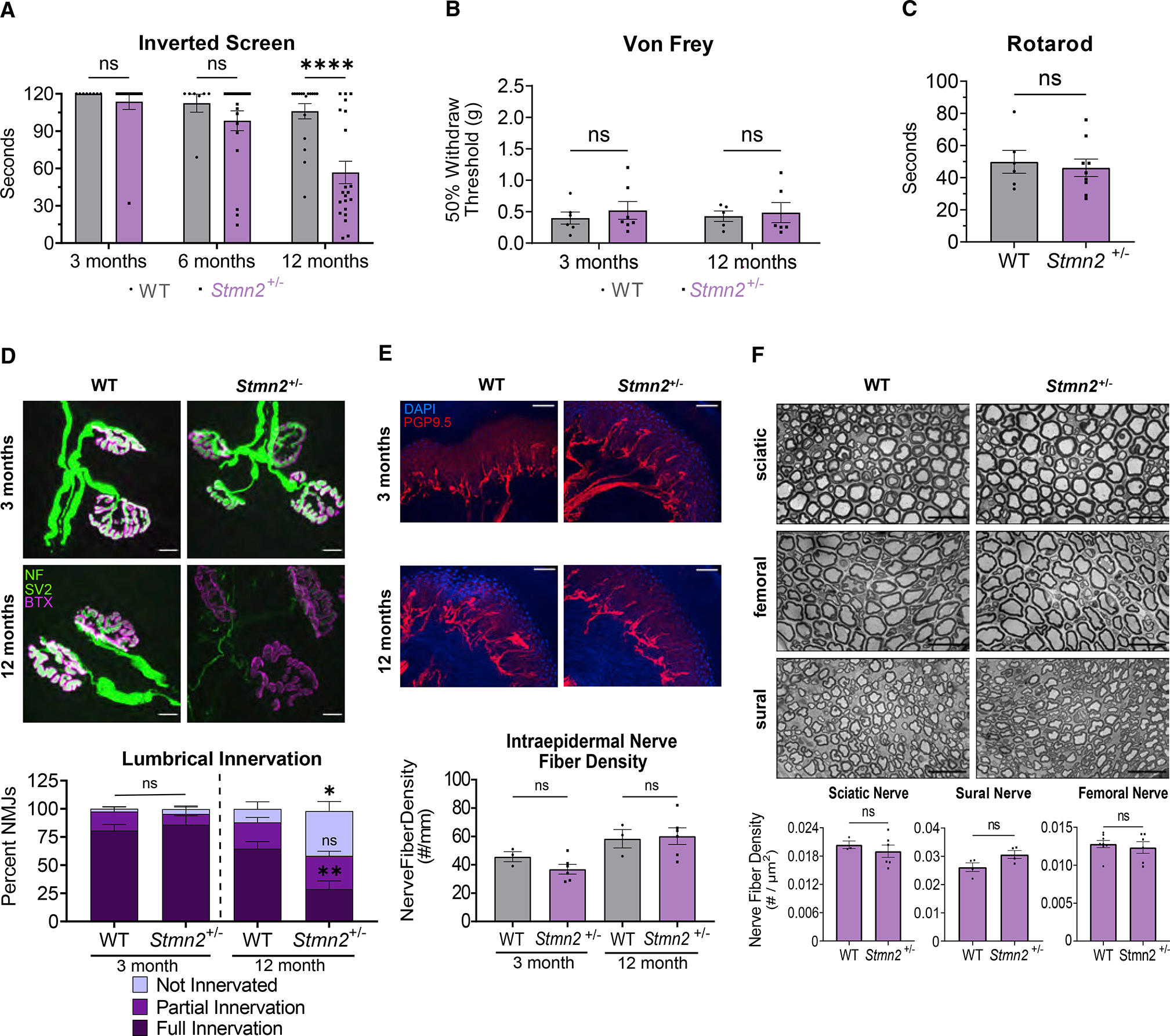
TDP-43 mediates proper Stathmin-2 (STMN2) mRNA splicing, and STMN2 protein is reduced in the spinal cord of most patients with amyotrophic lateral sclerosis (ALS). To test the hypothesis that STMN2 loss contributes to ALS pathogenesis, we generated constitutive and conditional STMN2 knockout mice. Constitutive STMN2 loss results in early-onset sensory and motor neuropathy featuring impaired motor behavior and dramatic distal neuromuscular junction (NMJ) denervation of fast-fatigable motor units, which are selectively vulnerable in ALS, without axon or motoneuron degeneration. Selective excision of STMN2 in motoneurons leads to similar NMJ pathology. STMN2 knockout heterozygous mice, which better model the partial loss of STMN2 protein found in patients with ALS, display a slowly progressive, motor-selective neuropathy with functional deficits and NMJ denervation. Thus, our findings strongly support the hypothesis that STMN2 reduction owing to TDP-43 pathology contributes to ALS pathogenesis.
Keywords: CP: Neuroscience; NMNAT2; SARM1; SCG-10; axon degeneration; motor neuron; neurodegeneration; neuropathy; stathmin.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.D. and J.M. are co-founders, scientific advisory board members, and shareholders of Disarm Therapeutics, a wholly owned subsidiary of Eli Lilly. A.J.B. is a consultant to Disarm Therapeutics. The authors have no other competing conflicts or financial interests.
Figures
Similar articles
-
Two cardinal features of ALS, reduced STMN2 and pathogenic TDP-43, synergize to accelerate motor decline in mice.Exp Neurol. 2025 Feb;384:115068. doi: 10.1016/j.expneurol.2024.115068. Epub 2024 Nov 26. Exp Neurol. 2025. PMID: 39603486
-
Loss of mouse Stmn2 function causes motor neuropathy.Neuron. 2022 May 18;110(10):1671-1688.e6. doi: 10.1016/j.neuron.2022.02.011. Epub 2022 Mar 15. Neuron. 2022. PMID: 35294901 Free PMC article.
-
ALS-implicated protein TDP-43 sustains levels of STMN2, a mediator of motor neuron growth and repair.Nat Neurosci. 2019 Feb;22(2):167-179. doi: 10.1038/s41593-018-0300-4. Epub 2019 Jan 14. Nat Neurosci. 2019. PMID: 30643292 Free PMC article.
-
Microtubules, Membranes, and Movement: New Roles for Stathmin-2 in Axon Integrity.J Neurosci Res. 2024 Sep;102(9):e25382. doi: 10.1002/jnr.25382. J Neurosci Res. 2024. PMID: 39253877 Free PMC article. Review.
-
TDP-43 dysregulation and neuromuscular junction disruption in amyotrophic lateral sclerosis.Transl Neurodegener. 2022 Dec 27;11(1):56. doi: 10.1186/s40035-022-00331-z. Transl Neurodegener. 2022. PMID: 36575535 Free PMC article. Review.
Cited by
-
Unveiling the double-edged sword: SOD1 trimers possess tissue-selective toxicity and bind septin-7 in motor neuron-like cells.Structure. 2024 Oct 3;32(10):1776-1792.e5. doi: 10.1016/j.str.2024.08.002. Epub 2024 Aug 28. Structure. 2024. PMID: 39208794
-
Synucleinopathy in Amyotrophic Lateral Sclerosis: A Potential Avenue for Antisense Therapeutics?Int J Mol Sci. 2022 Aug 19;23(16):9364. doi: 10.3390/ijms23169364. Int J Mol Sci. 2022. PMID: 36012622 Free PMC article. Review.
-
Antisense Oligonucleotides (ASOs) in Motor Neuron Diseases: A Road to Cure in Light and Shade.Int J Mol Sci. 2024 Apr 28;25(9):4809. doi: 10.3390/ijms25094809. Int J Mol Sci. 2024. PMID: 38732027 Free PMC article. Review.
-
RNA dysregulation in neurodegenerative diseases.EMBO J. 2025 Feb;44(3):613-638. doi: 10.1038/s44318-024-00352-6. Epub 2025 Jan 9. EMBO J. 2025. PMID: 39789319 Free PMC article. Review.
-
Alternative 3' UTR polyadenylation is disrupted in the rNLS8 mouse model of ALS/FTLD.Mol Brain. 2025 Jan 14;18(1):1. doi: 10.1186/s13041-025-01174-1. Mol Brain. 2025. PMID: 39810199 Free PMC article.
References
-
- Abbade J, Klemetti MM, Farrell A, Ermini L, Gillmore T, Sallais J, Tagliaferro A, Post M, and Caniggia I (2020). Increased placental mitochondrial fusion in gestational diabetes mellitus: an adaptive mechanism to optimize feto-placental metabolic homeostasis? BMJ Open Diabetes. Res Care. 8, e000923. 10.1136/bmjdrc-2019-000923. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
