

Selective enhancement of neural coding in V1 underlies fine-discrimination learning in tree shrew
- PMID: 35767997
- PMCID: PMC9378627
- DOI: 10.1016/j.cub.2022.06.009
Selective enhancement of neural coding in V1 underlies fine-discrimination learning in tree shrew
Abstract
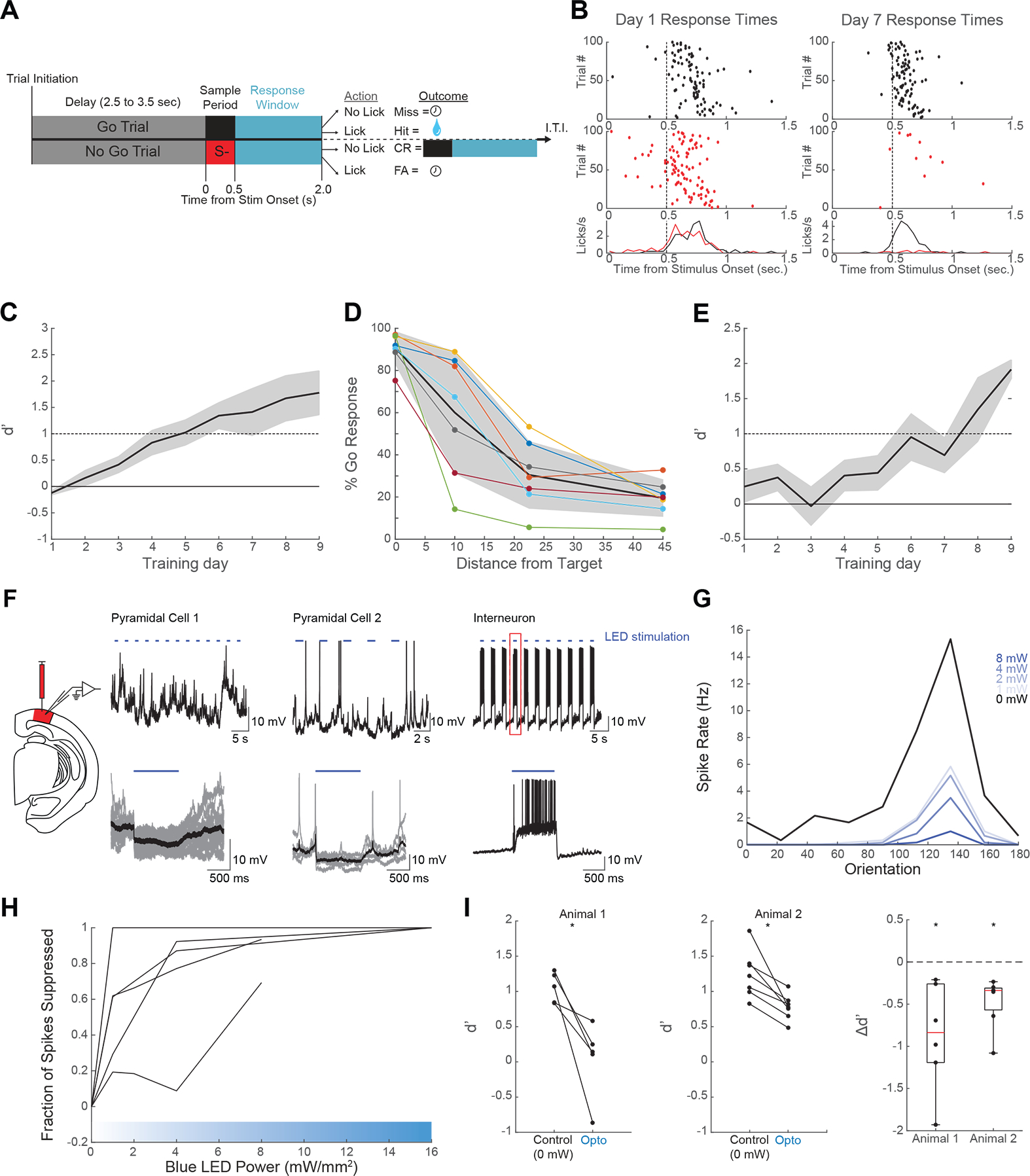
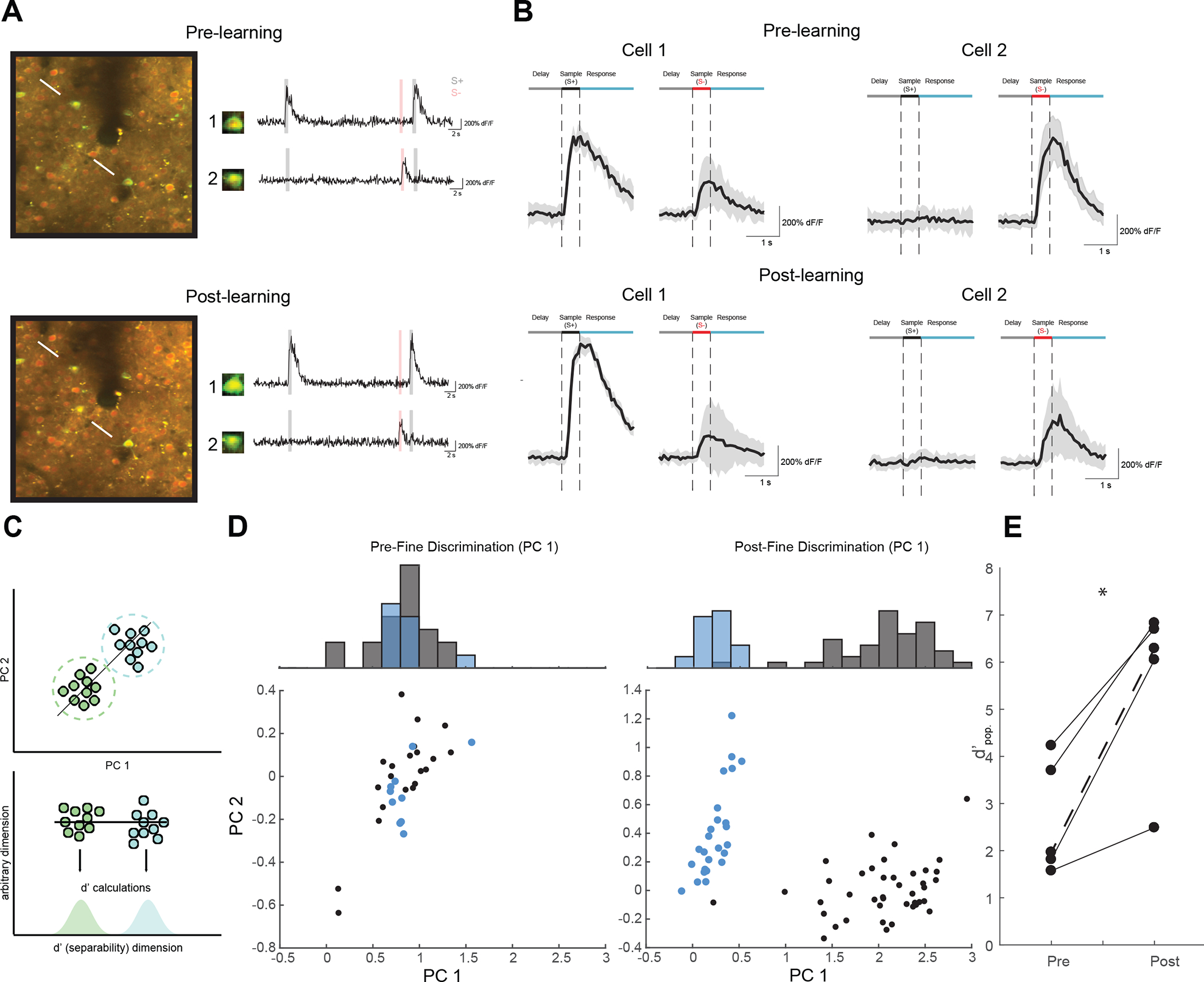
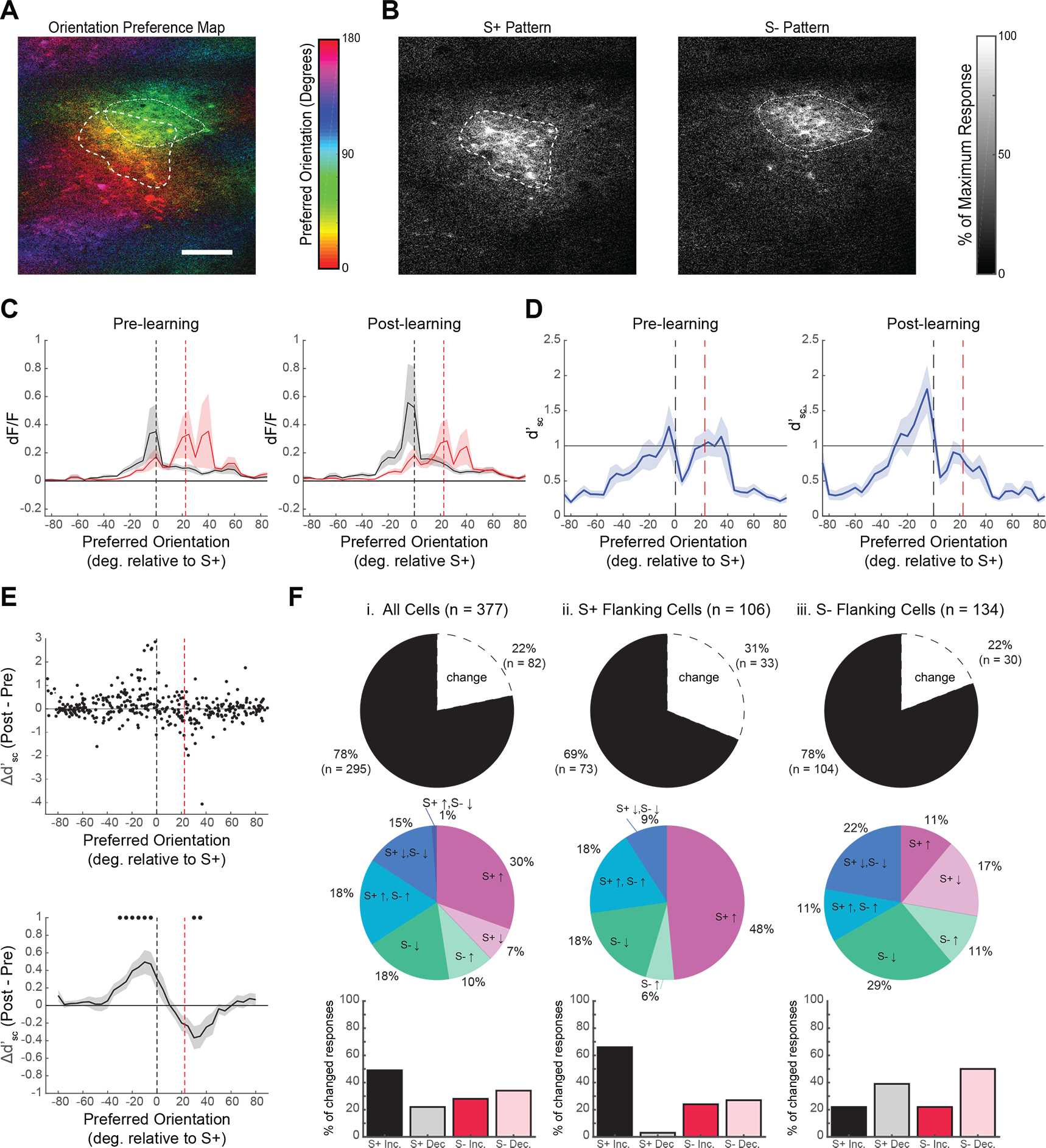
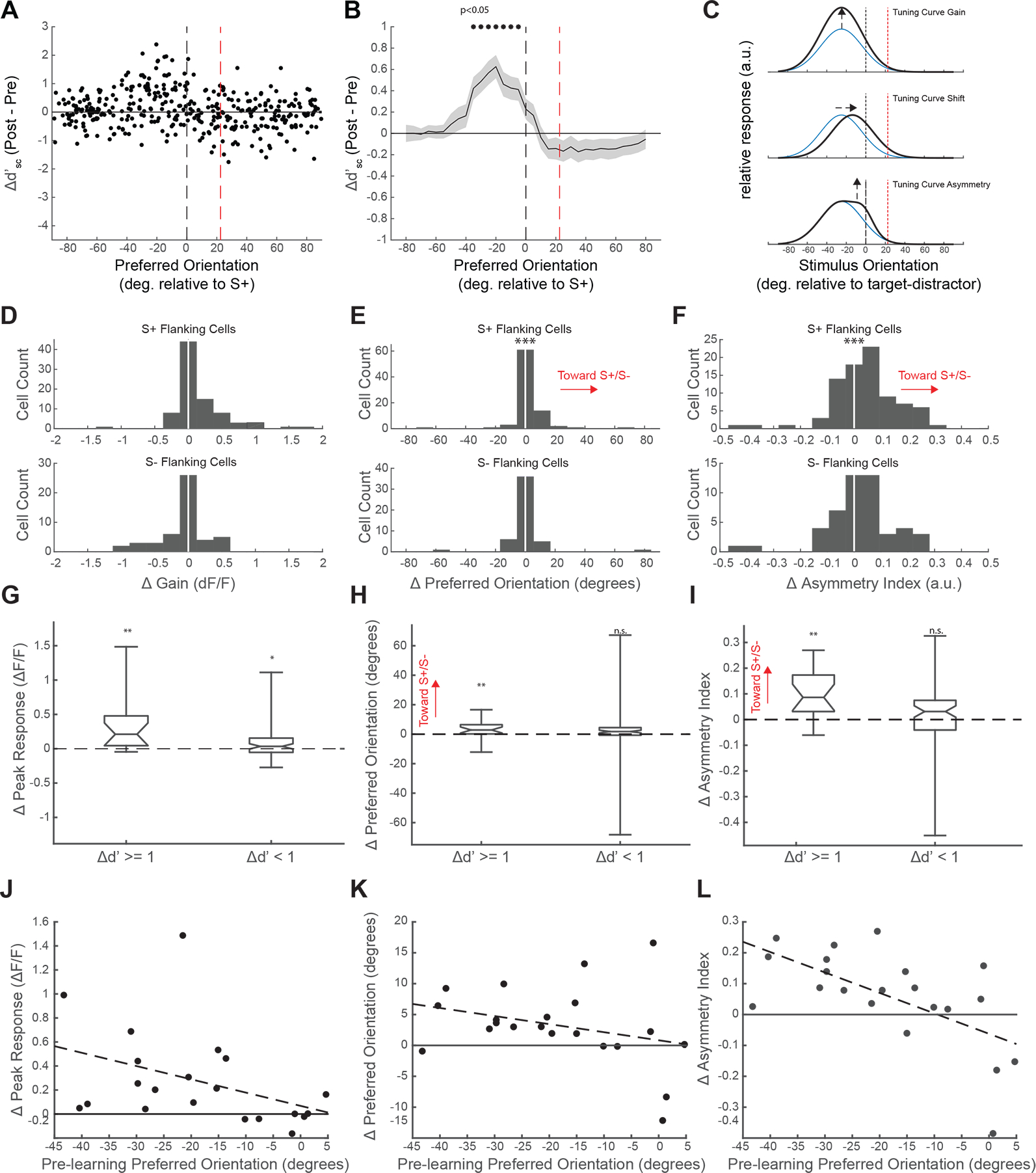
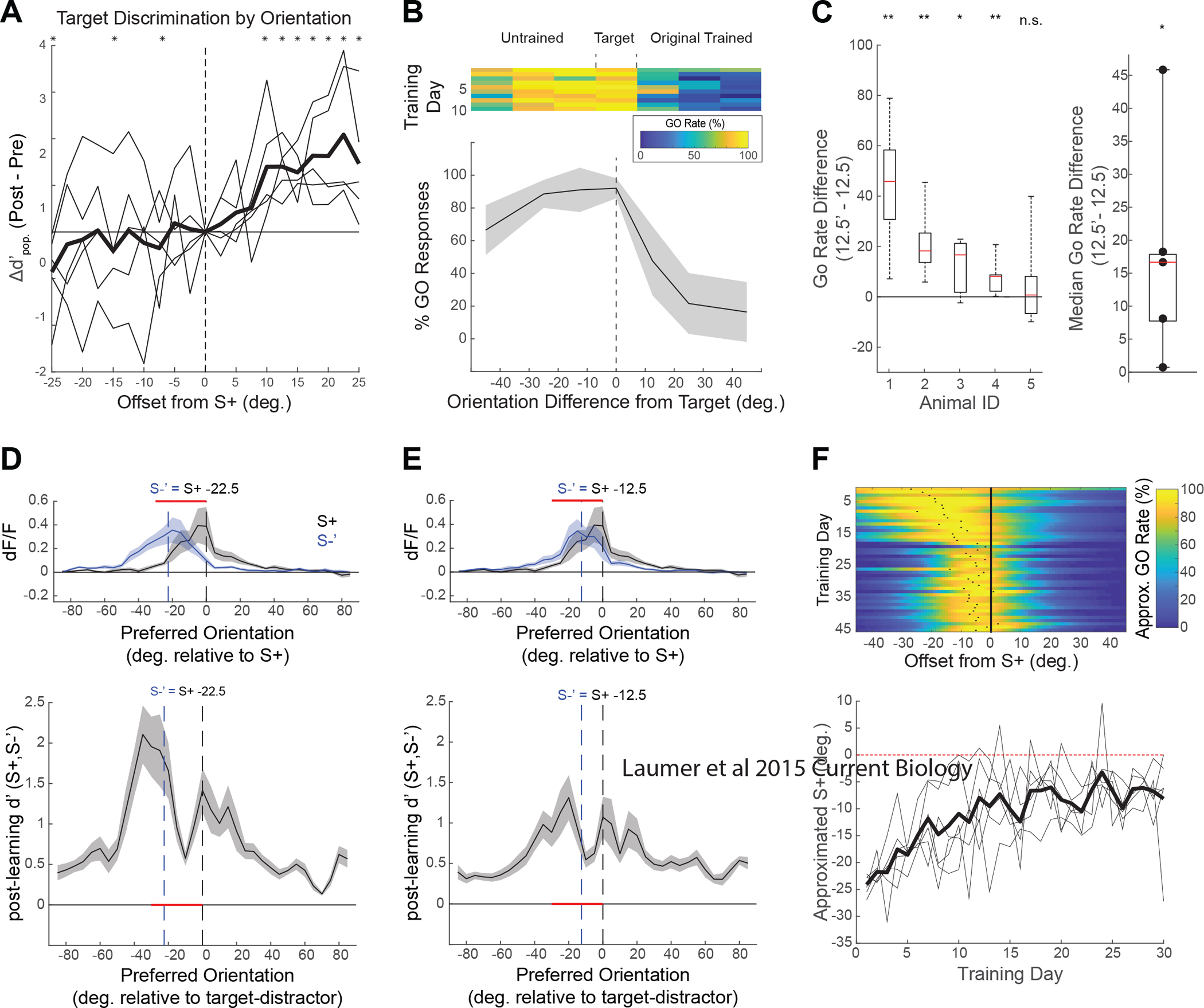
Visual discrimination improves with training, a phenomenon that is thought to reflect plastic changes in the responses of neurons in primary visual cortex (V1). However, the identity of the neurons that undergo change, the nature of the changes, and the consequences of these changes for other visual behaviors remain unclear. We used chronic in vivo 2-photon calcium imaging to monitor the responses of neurons in the V1 of tree shrews learning a Go/No-Go fine orientation discrimination task. We observed increases in neural population measures of discriminability for task-relevant stimuli that correlate with performance and depend on a select subset of neurons with preferred orientations that include the rewarded stimulus and nearby orientations biased away from the non-rewarded stimulus. Learning is accompanied by selective enhancement in the response of these neurons to the rewarded stimulus that further increases their ability to discriminate the task stimuli. These changes persist outside of the trained task and predict observed enhancement and impairment in performance of other discriminations, providing evidence for selective and persistent learning-induced plasticity in the V1, with significant consequences for perception.
Keywords: neural coding; neural discrimination; perceptual learning; tree shrew; visual cortex.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Visual neuroscience: A shrewd look at perceptual learning.Curr Biol. 2022 Aug 8;32(15):R839-R841. doi: 10.1016/j.cub.2022.07.002. Curr Biol. 2022. PMID: 35944484
Similar articles
-
Mechanism underpinning the sharpening of orientation and spatial frequency selectivities in the tree shrew (Tupaia belangeri) primary visual cortex.Brain Struct Funct. 2022 May;227(4):1265-1278. doi: 10.1007/s00429-021-02445-y. Epub 2022 Feb 3. Brain Struct Funct. 2022. PMID: 35118562
-
Mechanisms of visual perceptual learning in macaque visual cortex.Top Cogn Sci. 2010 Apr;2(2):239-50. doi: 10.1111/j.1756-8765.2009.01051.x. Epub 2009 Oct 30. Top Cogn Sci. 2010. PMID: 25163787
-
Learning Enhances Sensory Processing in Mouse V1 before Improving Behavior.J Neurosci. 2017 Jul 5;37(27):6460-6474. doi: 10.1523/JNEUROSCI.3485-16.2017. Epub 2017 May 30. J Neurosci. 2017. PMID: 28559381 Free PMC article.
-
Perceptual learning: is V1 up to the task?Curr Biol. 2004 Aug 24;14(16):R671-3. doi: 10.1016/j.cub.2004.08.016. Curr Biol. 2004. PMID: 15324688 Review.
-
Bottom-up and top-down dynamics in visual cortex.Prog Brain Res. 2005;149:65-81. doi: 10.1016/S0079-6123(05)49006-8. Prog Brain Res. 2005. PMID: 16226577 Review.
Cited by
-
Cellular Characterization and Interspecies Evolution of the Tree Shrew Retina across Postnatal Lifespan.Research (Wash D C). 2024 Nov 21;7:0536. doi: 10.34133/research.0536. eCollection 2024. Research (Wash D C). 2024. PMID: 39574940 Free PMC article.
-
Cognitive Deficits and Alzheimer's Disease-Like Pathologies in the Aged Chinese Tree Shrew.Mol Neurobiol. 2024 Apr;61(4):1892-1906. doi: 10.1007/s12035-023-03663-7. Epub 2023 Oct 9. Mol Neurobiol. 2024. PMID: 37814108
-
High-throughput markerless pose estimation and home-cage activity analysis of tree shrew using deep learning.Animal Model Exp Med. 2025 May;8(5):896-905. doi: 10.1002/ame2.12530. Epub 2025 Jan 23. Animal Model Exp Med. 2025. PMID: 39846430 Free PMC article.
-
Neural circuits for binocular vision: Ocular dominance, interocular matching, and disparity selectivity.Front Neural Circuits. 2023 Feb 15;17:1084027. doi: 10.3389/fncir.2023.1084027. eCollection 2023. Front Neural Circuits. 2023. PMID: 36874946 Free PMC article. Review.
-
A compressed hierarchy for visual form processing in the tree shrew.Nature. 2025 Aug 27. doi: 10.1038/s41586-025-09441-w. Online ahead of print. Nature. 2025. PMID: 40866712
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
