

Structural and electrophysiological basis for the modulation of KCNQ1 channel currents by ML277
- PMID: 35768468
- PMCID: PMC9243137
- DOI: 10.1038/s41467-022-31526-7
Structural and electrophysiological basis for the modulation of KCNQ1 channel currents by ML277
Abstract
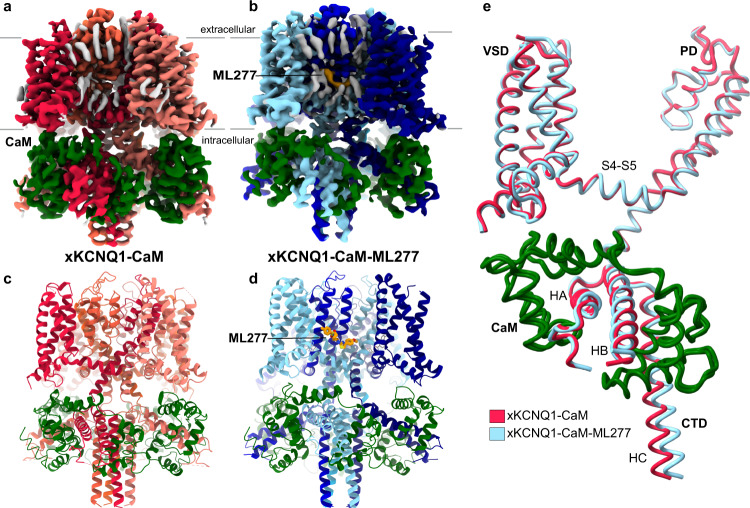
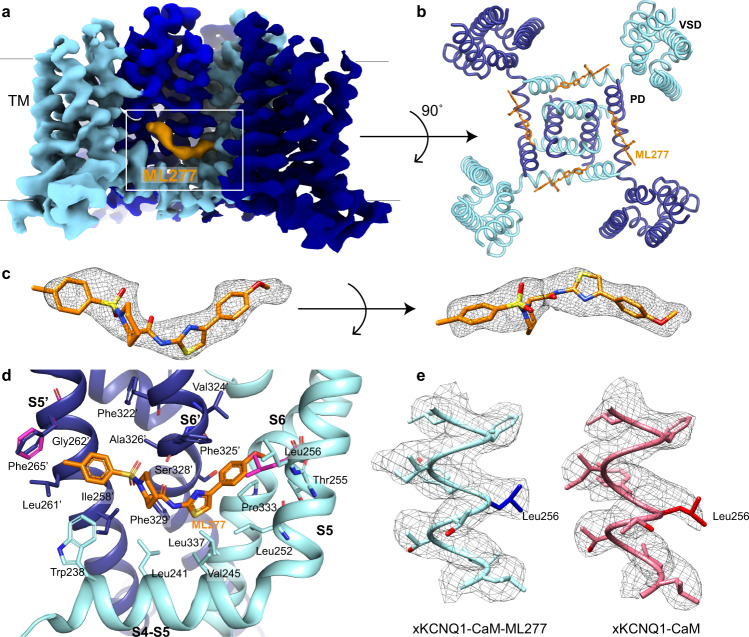
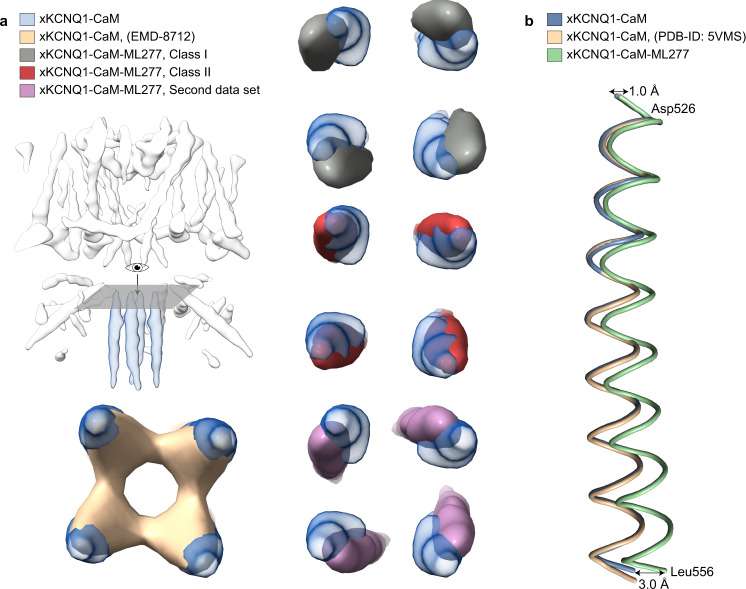
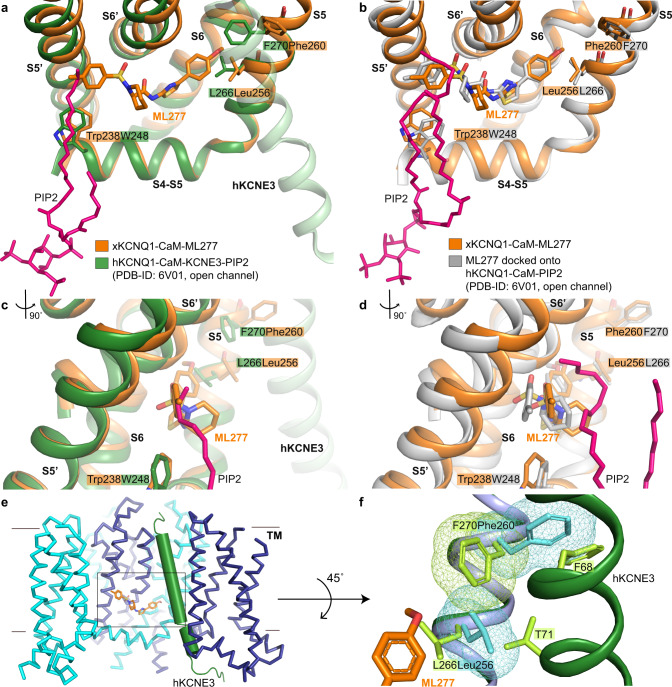
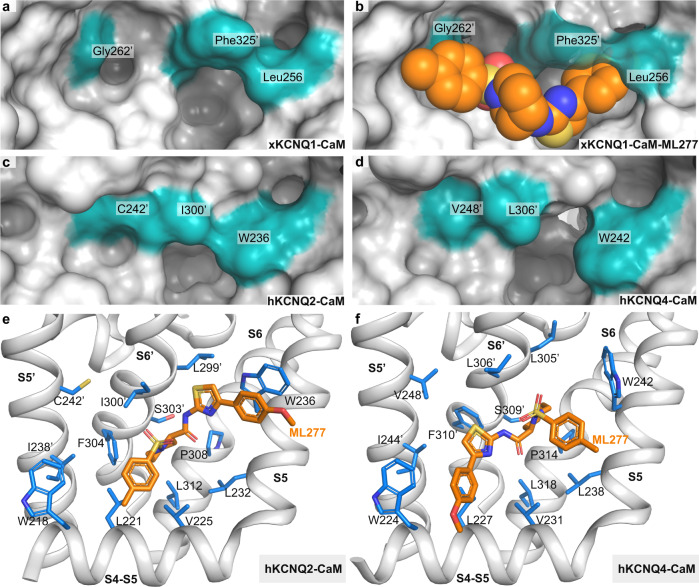
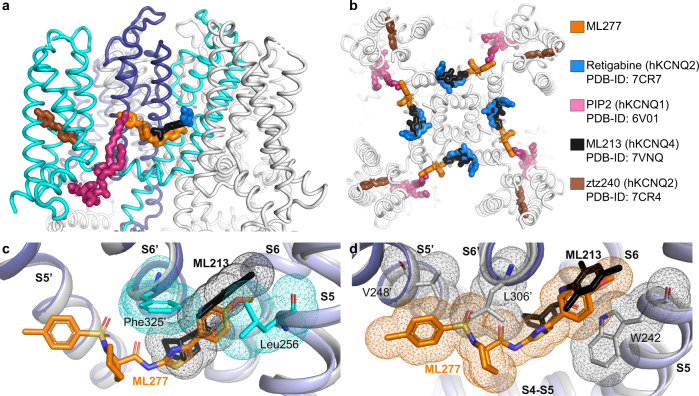
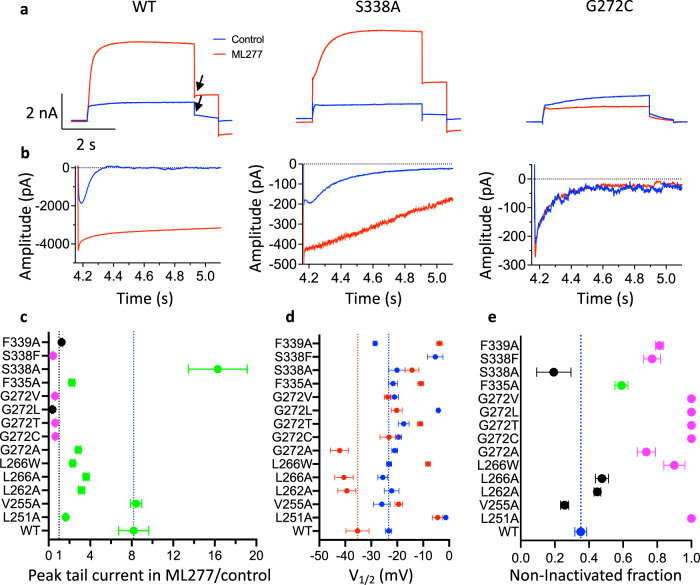
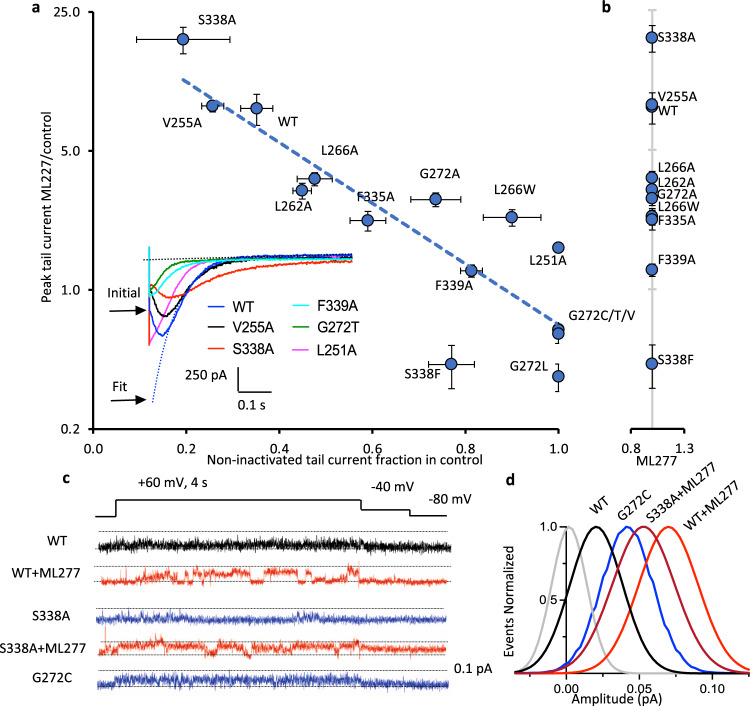
The KCNQ1 ion channel plays critical physiological roles in electrical excitability and K+ recycling in organs including the heart, brain, and gut. Loss of function is relatively common and can cause sudden arrhythmic death, sudden infant death, epilepsy and deafness. Here, we report cryogenic electron microscopic (cryo-EM) structures of Xenopus KCNQ1 bound to Ca2+/Calmodulin, with and without the KCNQ1 channel activator, ML277. A single binding site for ML277 was identified, localized to a pocket lined by the S4-S5 linker, S5 and S6 helices of two separate subunits. Several pocket residues are not conserved in other KCNQ isoforms, explaining specificity. MD simulations and point mutations support this binding location for ML277 in open and closed channels and reveal that prevention of inactivation is an important component of the activator effect. Our work provides direction for therapeutic intervention targeting KCNQ1 loss of function pathologies including long QT interval syndrome and seizures.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
ML277 specifically enhances the fully activated open state of KCNQ1 by modulating VSD-pore coupling.Elife. 2019 Jul 22;8:e48576. doi: 10.7554/eLife.48576. Elife. 2019. PMID: 31329101 Free PMC article.
-
Dynamic subunit stoichiometry confers a progressive continuum of pharmacological sensitivity by KCNQ potassium channels.Proc Natl Acad Sci U S A. 2013 May 21;110(21):8732-7. doi: 10.1073/pnas.1300684110. Epub 2013 May 6. Proc Natl Acad Sci U S A. 2013. PMID: 23650380 Free PMC article.
-
Identification of (R)-N-(4-(4-methoxyphenyl)thiazol-2-yl)-1-tosylpiperidine-2-carboxamide, ML277, as a novel, potent and selective K(v)7.1 (KCNQ1) potassium channel activator.Bioorg Med Chem Lett. 2012 Sep 15;22(18):5936-41. doi: 10.1016/j.bmcl.2012.07.060. Epub 2012 Aug 2. Bioorg Med Chem Lett. 2012. PMID: 22910039 Free PMC article.
-
Insights into Cardiac IKs (KCNQ1/KCNE1) Channels Regulation.Int J Mol Sci. 2020 Dec 11;21(24):9440. doi: 10.3390/ijms21249440. Int J Mol Sci. 2020. PMID: 33322401 Free PMC article. Review.
-
The membrane protein KCNQ1 potassium ion channel: Functional diversity and current structural insights.Biochim Biophys Acta Biomembr. 2020 May 1;1862(5):183148. doi: 10.1016/j.bbamem.2019.183148. Epub 2019 Dec 9. Biochim Biophys Acta Biomembr. 2020. PMID: 31825788 Free PMC article. Review.
Cited by
-
Constitutive opening of the Kv7.2 pore activation gate causes KCNQ2-developmental encephalopathy.Proc Natl Acad Sci U S A. 2024 Dec 3;121(49):e2412388121. doi: 10.1073/pnas.2412388121. Epub 2024 Nov 27. Proc Natl Acad Sci U S A. 2024. PMID: 39602259 Free PMC article.
-
Atypical KCNQ1/Kv7 channel function in a neonatal diabetes patient: Hypersecretion preceded the failure of pancreatic β-cells.iScience. 2024 Jun 17;27(7):110291. doi: 10.1016/j.isci.2024.110291. eCollection 2024 Jul 19. iScience. 2024. PMID: 39055936 Free PMC article.
-
Mechanisms of KCNQ1 gating modulation by KCNE1/3 for cell-specific function.Cell Res. 2025 Jul 31. doi: 10.1038/s41422-025-01152-1. Online ahead of print. Cell Res. 2025. PMID: 40745202
-
A generic binding pocket for small molecule IKs activators at the extracellular inter-subunit interface of KCNQ1 and KCNE1 channel complexes.Elife. 2023 Sep 14;12:RP87038. doi: 10.7554/eLife.87038. Elife. 2023. PMID: 37707495 Free PMC article.
-
In vitro human cell culture models in a bench-to-bedside approach to epilepsy.Epilepsia Open. 2024 Jun;9(3):865-890. doi: 10.1002/epi4.12941. Epub 2024 Apr 18. Epilepsia Open. 2024. PMID: 38637998 Free PMC article. Review.
References
-
- Sanguinetti, M. C. & Seebohm, G. Physiological functions, biophysical properties, and regulation of KCNQ1 (KV7.1) potassium channels. Adv. Exp. Med. Biol.1349, 335–353 (2021). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
