

Unifying turbulent dynamics framework distinguishes different brain states
- PMID: 35768641
- PMCID: PMC9243255
- DOI: 10.1038/s42003-022-03576-6
Unifying turbulent dynamics framework distinguishes different brain states
Abstract
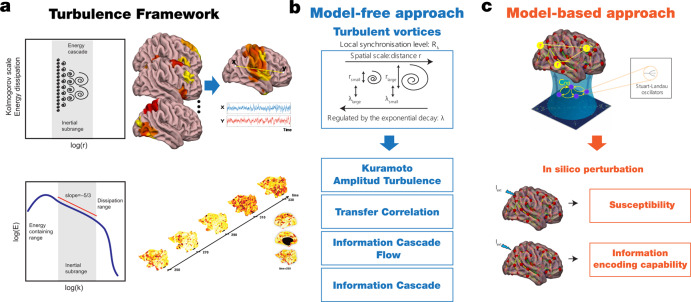
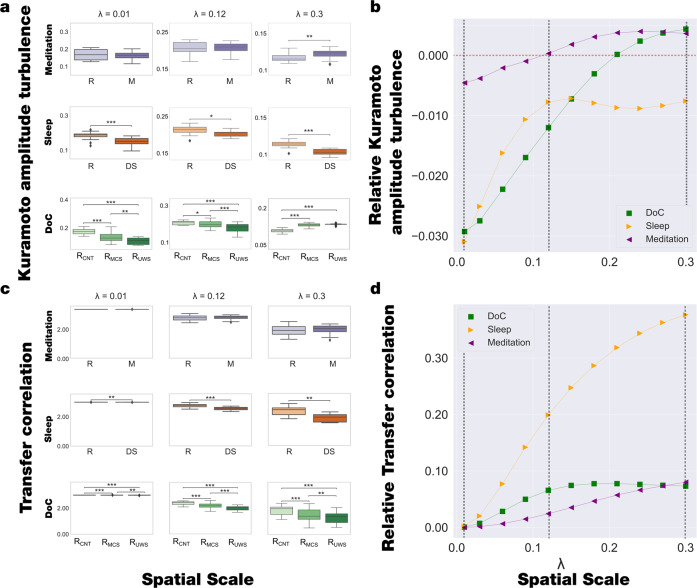
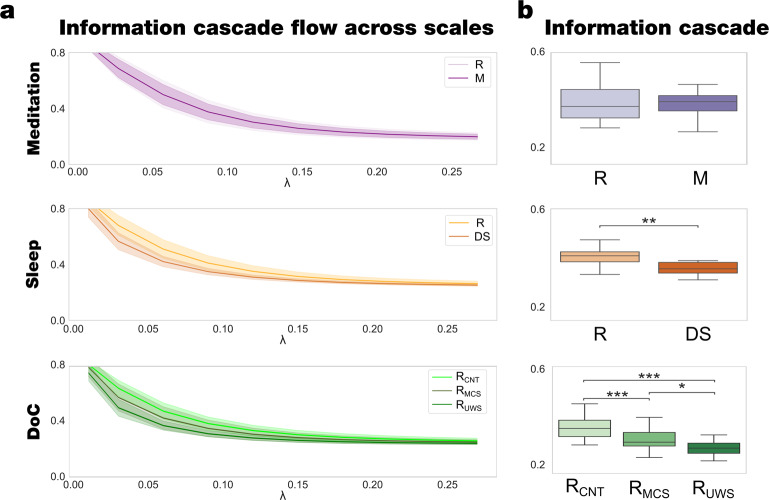
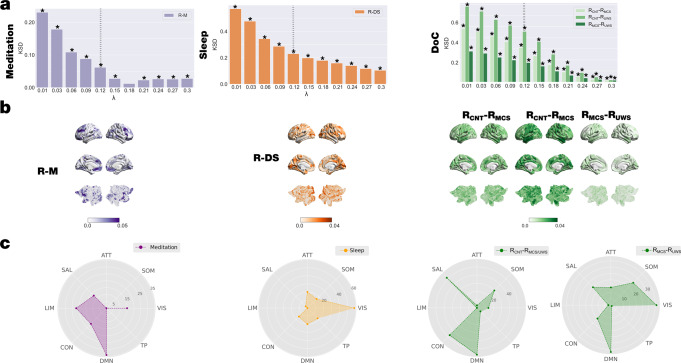
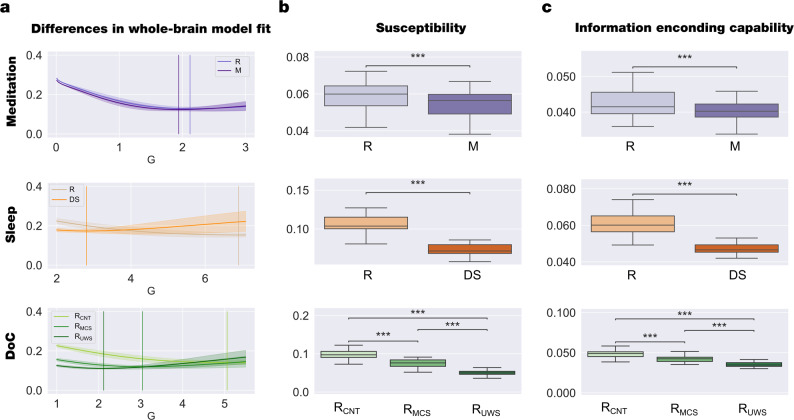
Significant advances have been made by identifying the levels of synchrony of the underlying dynamics of a given brain state. This research has demonstrated that non-conscious dynamics tend to be more synchronous than in conscious states, which are more asynchronous. Here we go beyond this dichotomy to demonstrate that different brain states are underpinned by dissociable spatiotemporal dynamics. We investigated human neuroimaging data from different brain states (resting state, meditation, deep sleep and disorders of consciousness after coma). The model-free approach was based on Kuramoto's turbulence framework using coupled oscillators. This was extended by a measure of the information cascade across spatial scales. Complementarily, the model-based approach used exhaustive in silico perturbations of whole-brain models fitted to these measures. This allowed studying of the information encoding capabilities in given brain states. Overall, this framework demonstrates that elements from turbulence theory provide excellent tools for describing and differentiating between brain states.
© 2022. The Author(s).
Conflict of interest statement
E.T. is an Editorial Board Member for
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
