

The mTORC1-SLC4A7 axis stimulates bicarbonate import to enhance de novo nucleotide synthesis
- PMID: 35772404
- PMCID: PMC9444906
- DOI: 10.1016/j.molcel.2022.06.008
The mTORC1-SLC4A7 axis stimulates bicarbonate import to enhance de novo nucleotide synthesis
Abstract
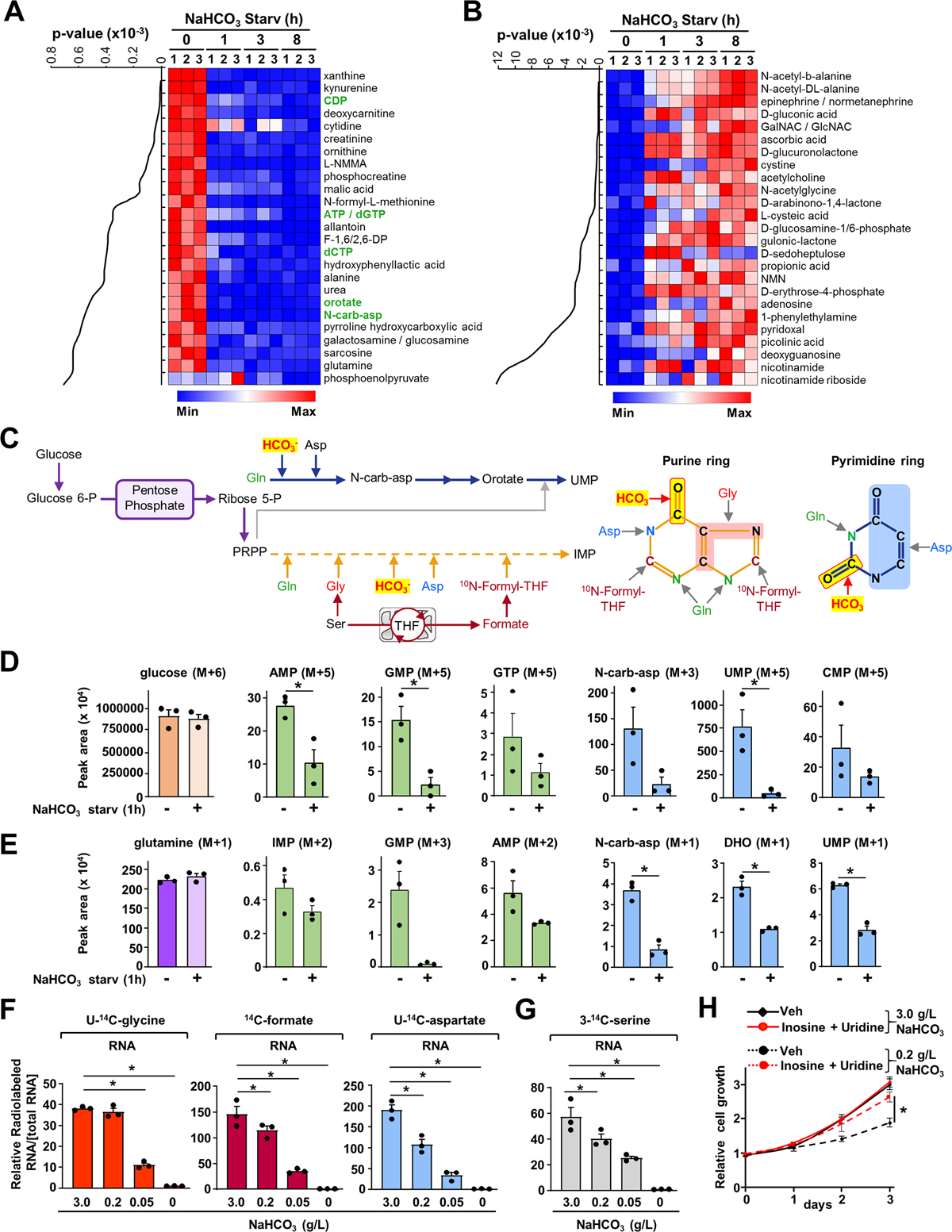
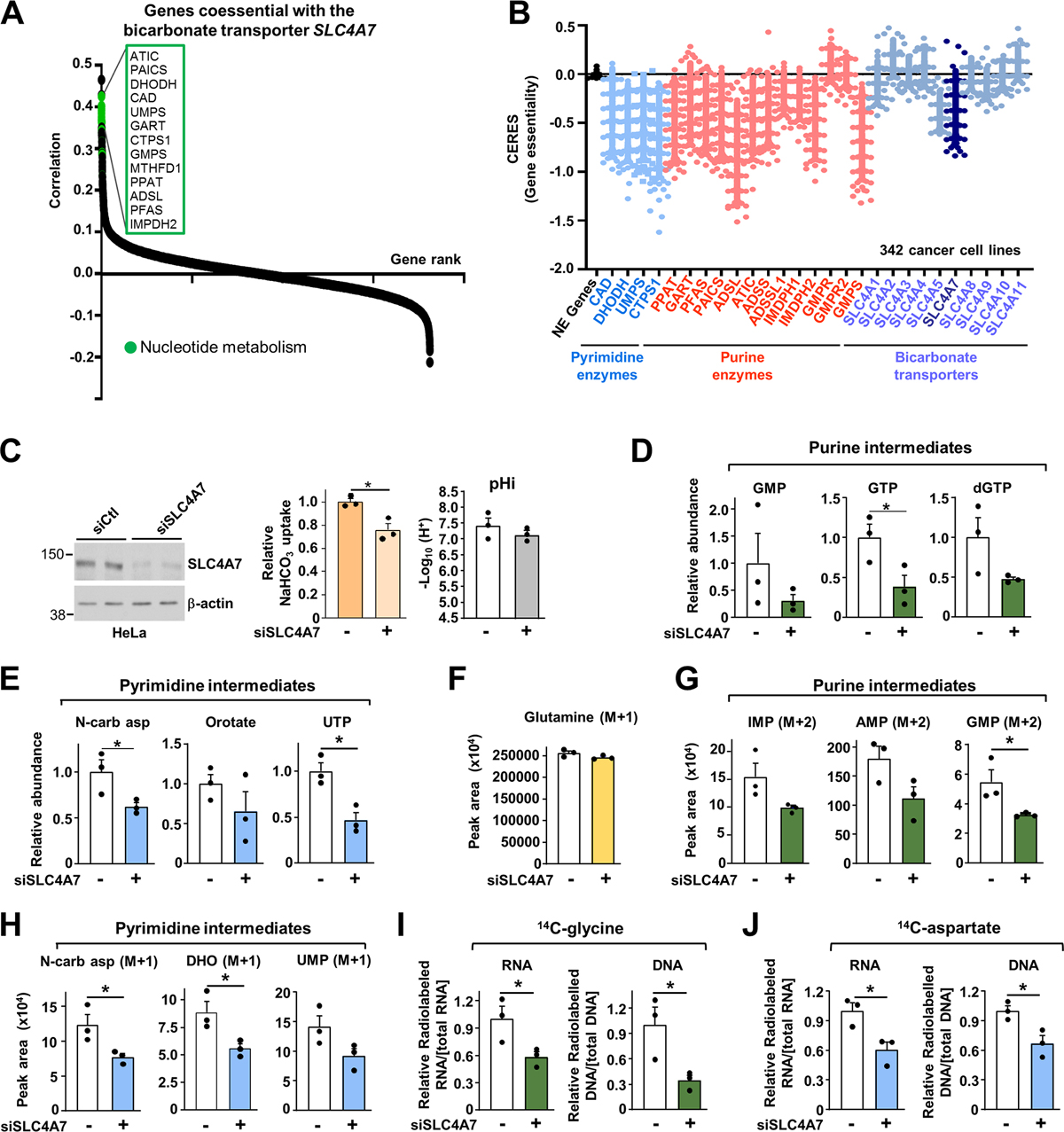
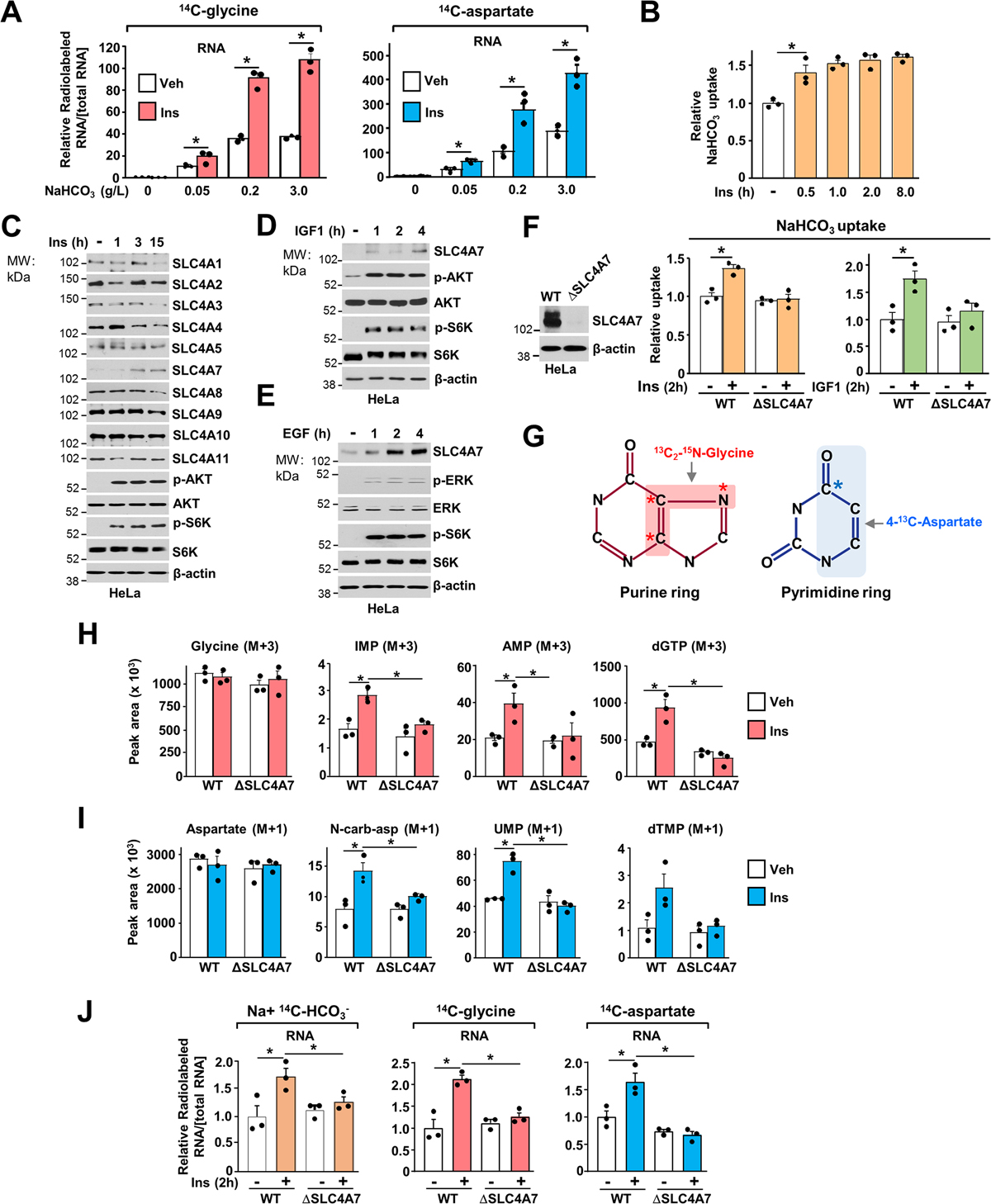
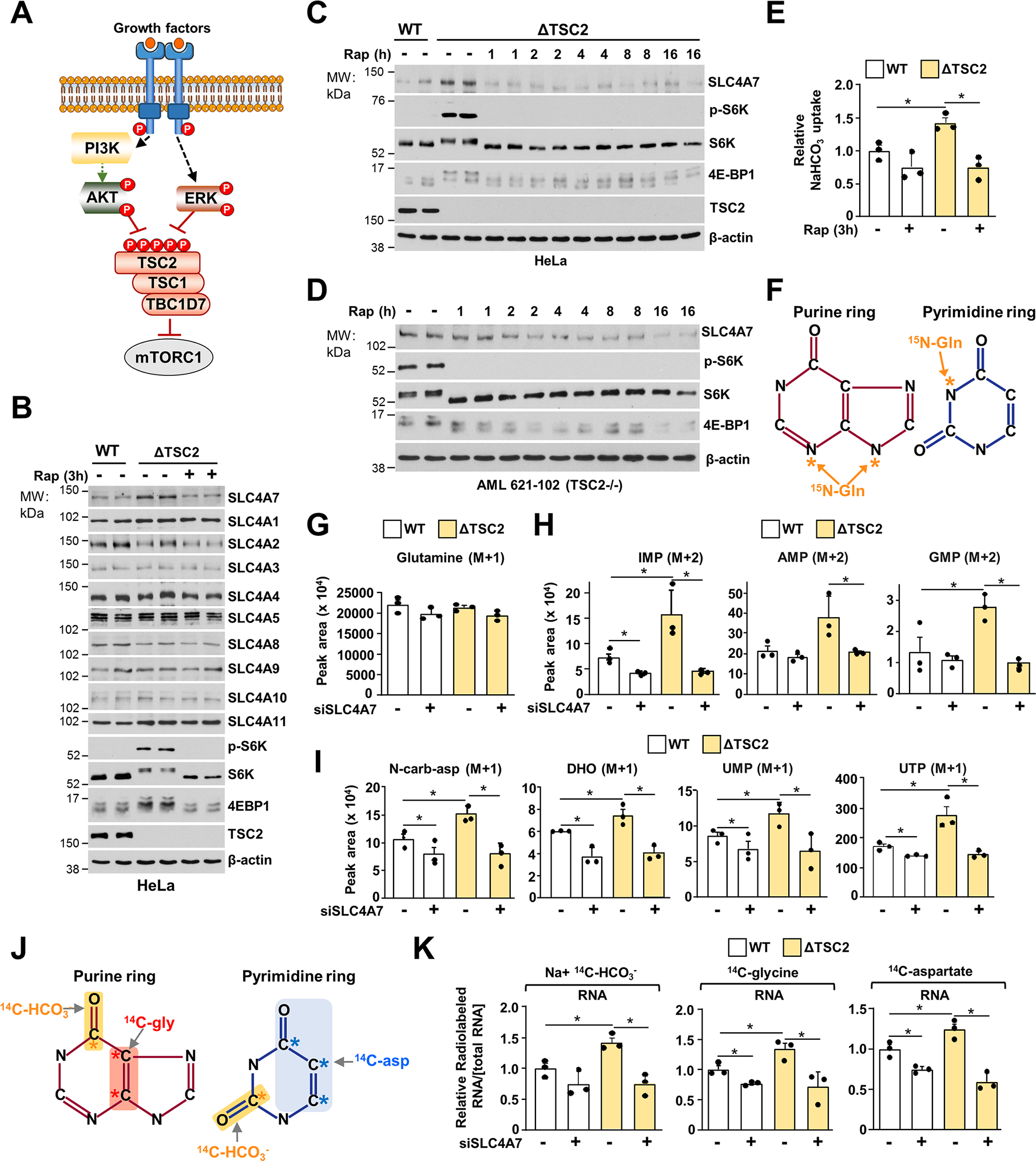
Bicarbonate (HCO3-) ions maintain pH homeostasis in eukaryotic cells and serve as a carbonyl donor to support cellular metabolism. However, whether the abundance of HCO3- is regulated or harnessed to promote cell growth is unknown. The mechanistic target of rapamycin complex 1 (mTORC1) adjusts cellular metabolism to support biomass production and cell growth. We find that mTORC1 stimulates the intracellular transport of HCO3- to promote nucleotide synthesis through the selective translational regulation of the sodium bicarbonate cotransporter SLC4A7. Downstream of mTORC1, SLC4A7 mRNA translation required the S6K-dependent phosphorylation of the translation factor eIF4B. In mTORC1-driven cells, loss of SLC4A7 resulted in reduced cell and tumor growth and decreased flux through de novo purine and pyrimidine synthesis in human cells and tumors without altering the intracellular pH. Thus, mTORC1 signaling, through the control of SLC4A7 expression, harnesses environmental bicarbonate to promote anabolic metabolism, cell biomass, and growth.
Keywords: SLC4A7/NBCn1; bicarbonate metabolism; mTOR signaling; purine metabolism; pyrimidine metabolism.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflict of interests.
Figures
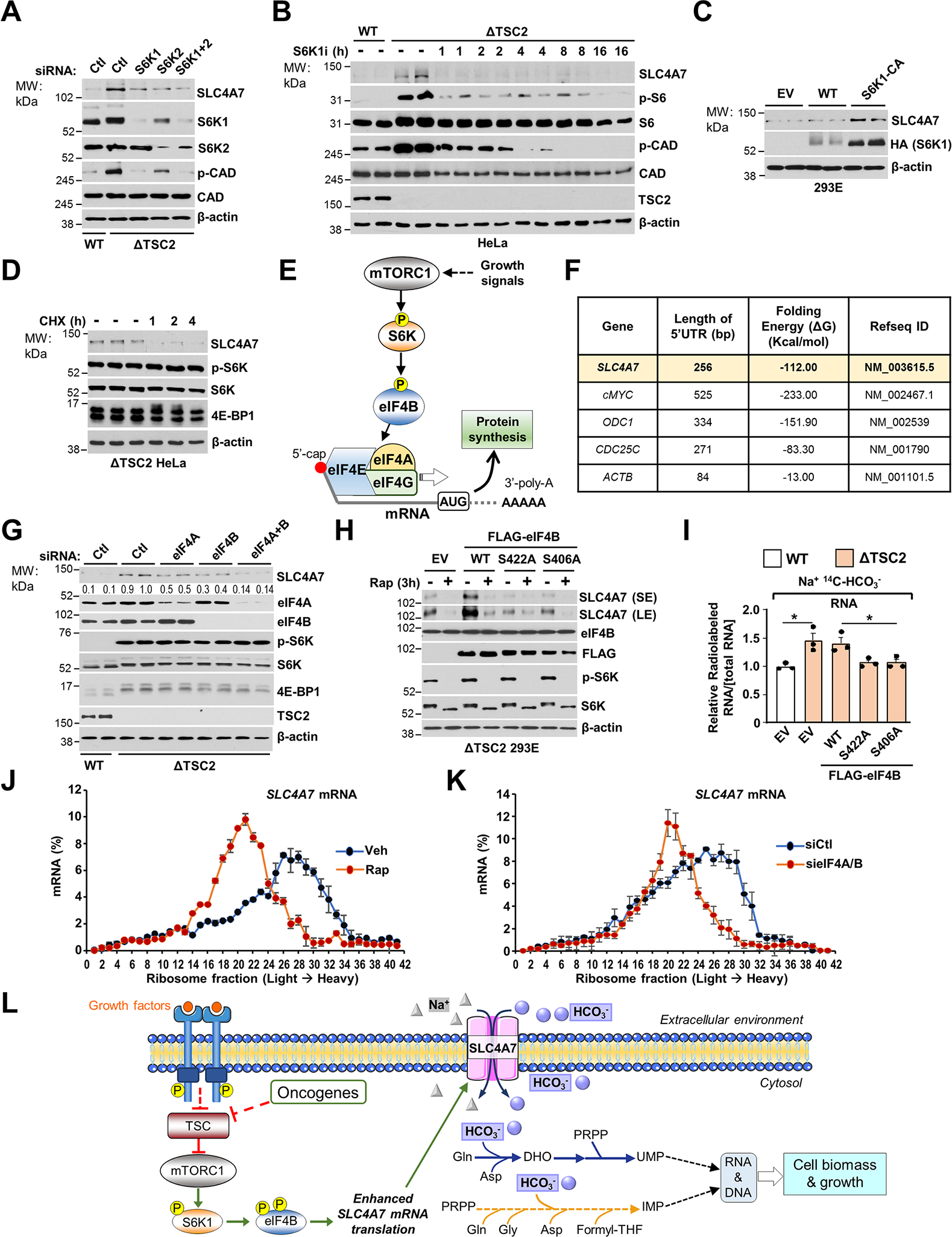
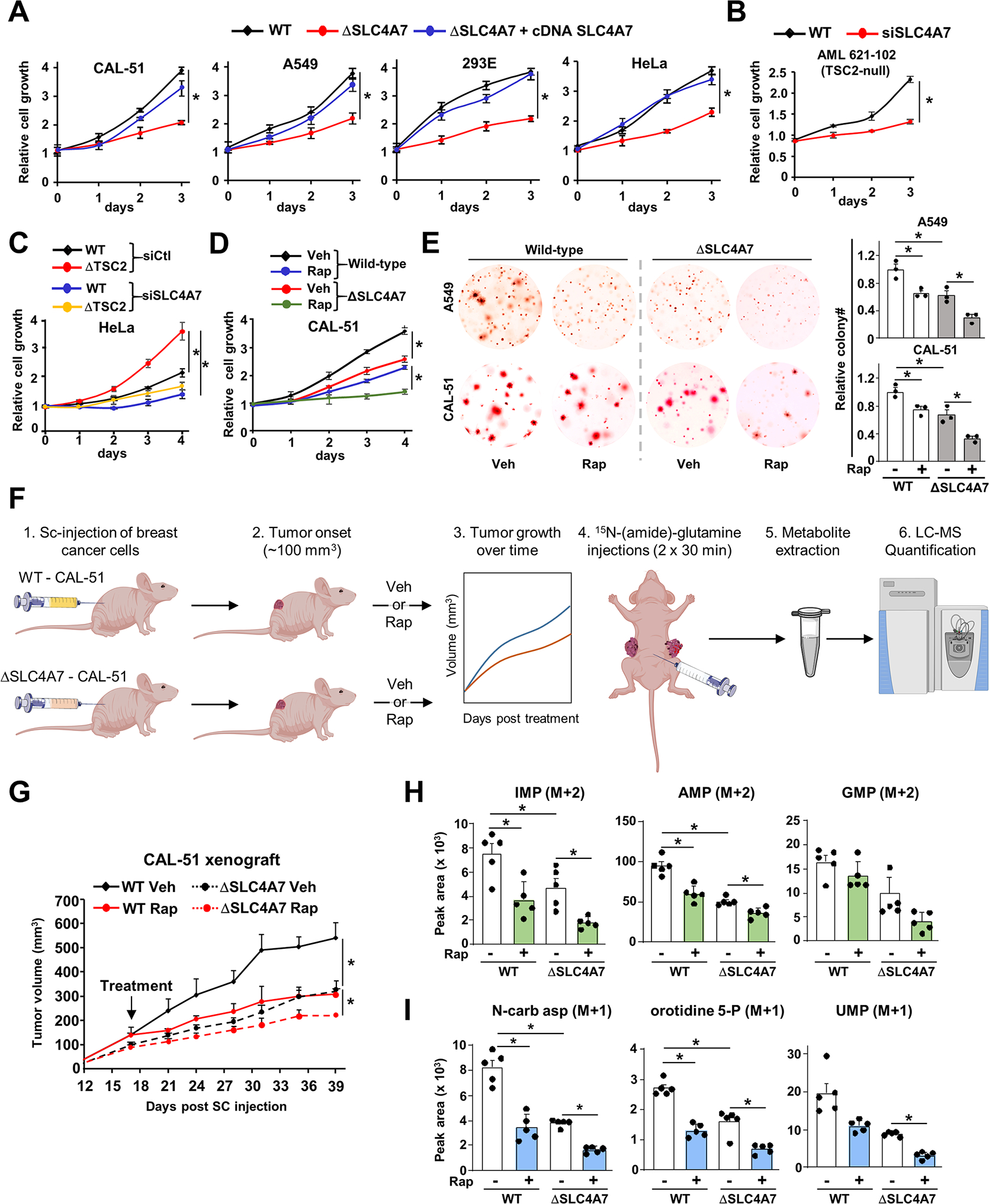
Comment in
-
SLC4A7 and mTORC1 raise nucleotide synthesis with bicarbonate.Mol Cell. 2022 Sep 1;82(17):3121-3123. doi: 10.1016/j.molcel.2022.08.010. Mol Cell. 2022. PMID: 36055205
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
