

Antibody escape of SARS-CoV-2 Omicron BA.4 and BA.5 from vaccine and BA.1 serum
- PMID: 35772405
- PMCID: PMC9181312
- DOI: 10.1016/j.cell.2022.06.005
Antibody escape of SARS-CoV-2 Omicron BA.4 and BA.5 from vaccine and BA.1 serum
Abstract
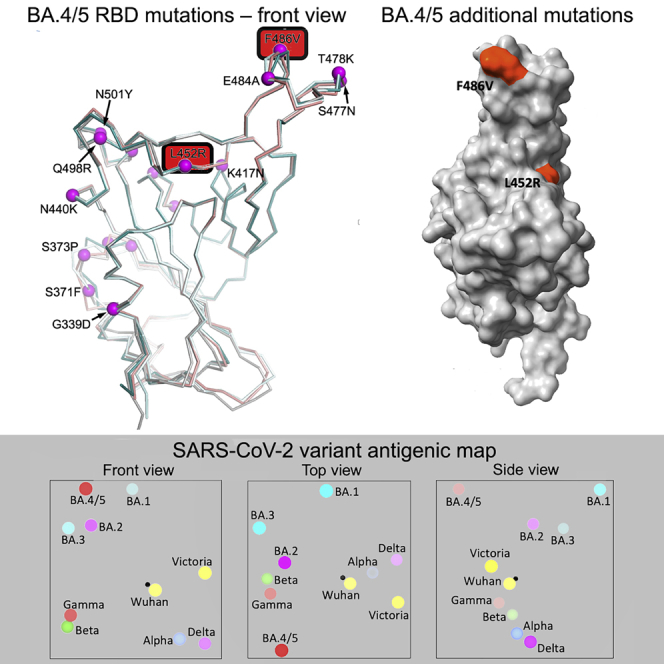
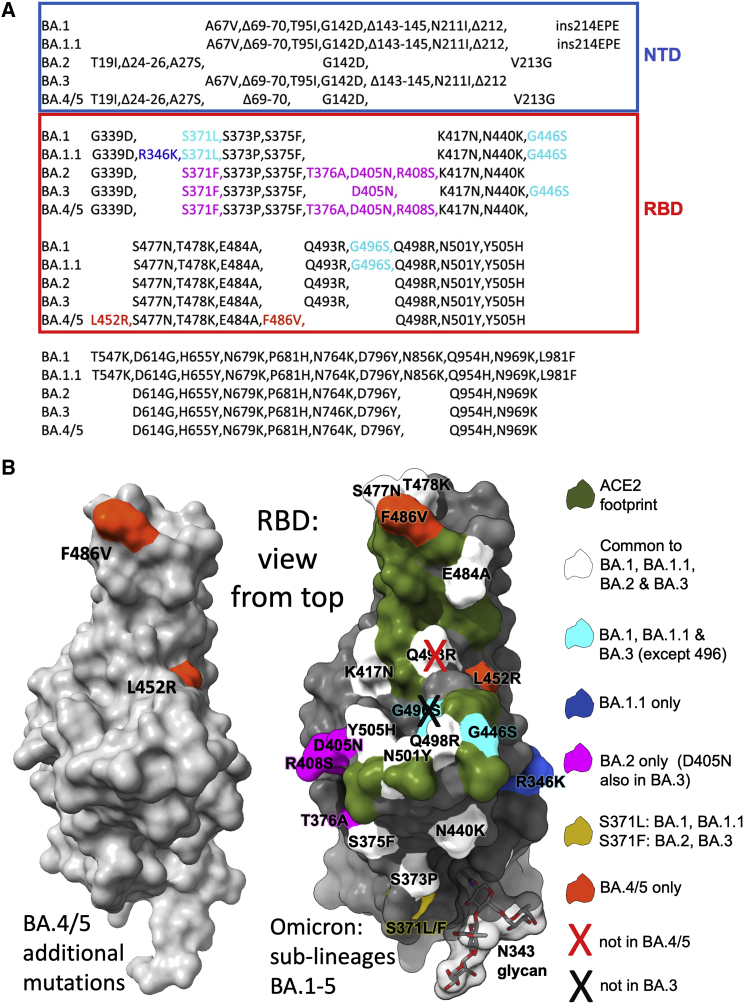
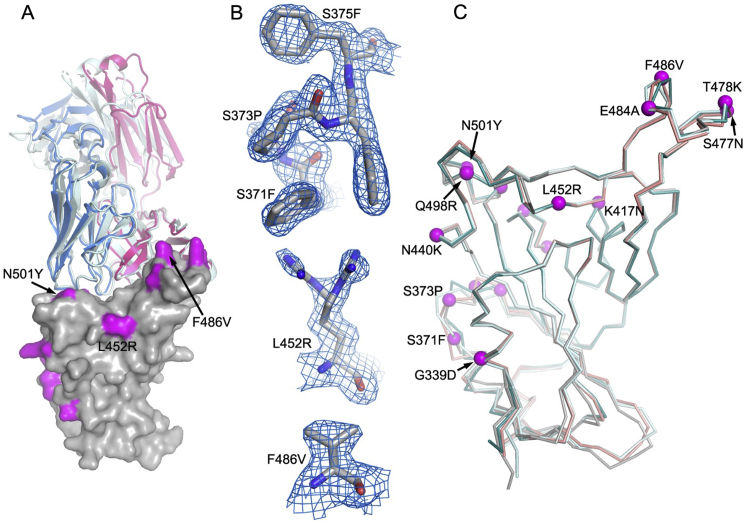
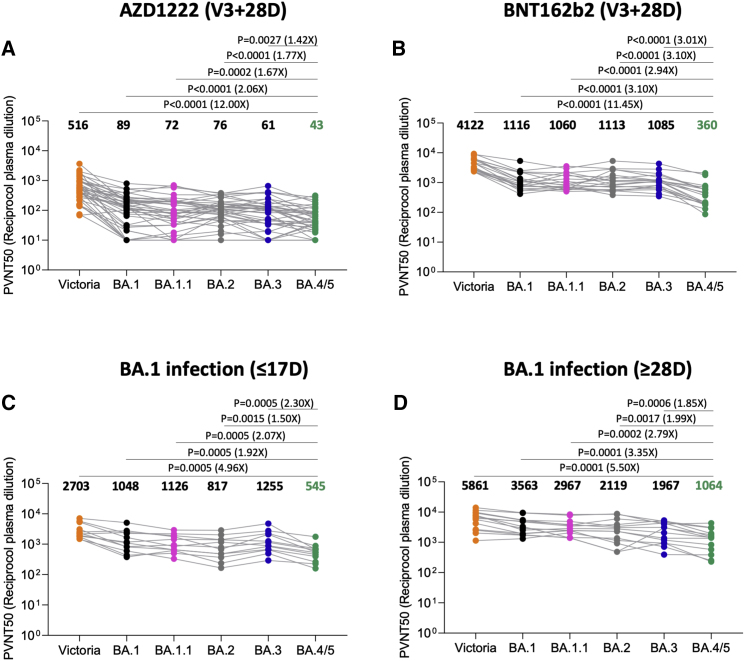
The Omicron lineage of SARS-CoV-2, which was first described in November 2021, spread rapidly to become globally dominant and has split into a number of sublineages. BA.1 dominated the initial wave but has been replaced by BA.2 in many countries. Recent sequencing from South Africa's Gauteng region uncovered two new sublineages, BA.4 and BA.5, which are taking over locally, driving a new wave. BA.4 and BA.5 contain identical spike sequences, and although closely related to BA.2, they contain further mutations in the receptor-binding domain of their spikes. Here, we study the neutralization of BA.4/5 using a range of vaccine and naturally immune serum and panels of monoclonal antibodies. BA.4/5 shows reduced neutralization by the serum from individuals vaccinated with triple doses of AstraZeneca or Pfizer vaccine compared with BA.1 and BA.2. Furthermore, using the serum from BA.1 vaccine breakthrough infections, there are, likewise, significant reductions in the neutralization of BA.4/5, raising the possibility of repeat Omicron infections.
Keywords: BA.4; BA.5; COVID-19; Omicron; SARS-CoV-2; VoC; antibody escape; variant.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.R.S. sits on the GSK Vaccines Scientific Advisory Board and is a founding member of RQ Biotechnology. Oxford University holds intellectual property related to the Oxford-AstraZeneca vaccine and SARS-CoV-2 mAb discovered in G.R.S.’s laboratory. A.J.P. is Chair of UK Dept. health and Social Care’s (DHSC) Joint Committee on Vaccination & Immunisation (JCVI) but does not participate in the JCVI COVID-19 committee and is a member of the WHO’s SAGE. The views expressed in this article do not necessarily represent the views of DHSC, JCVI, or WHO. The University of Oxford has entered into a partnership with AstraZeneca on coronavirus vaccine development. T.L. is named as an inventor on a patent application covering this SARS-CoV-2 vaccine and was a consultant to Vaccitech for an unrelated project while the study was conducted. S.J.D. is a scientific advisor to the Scottish Parliament on COVID-19.
Figures
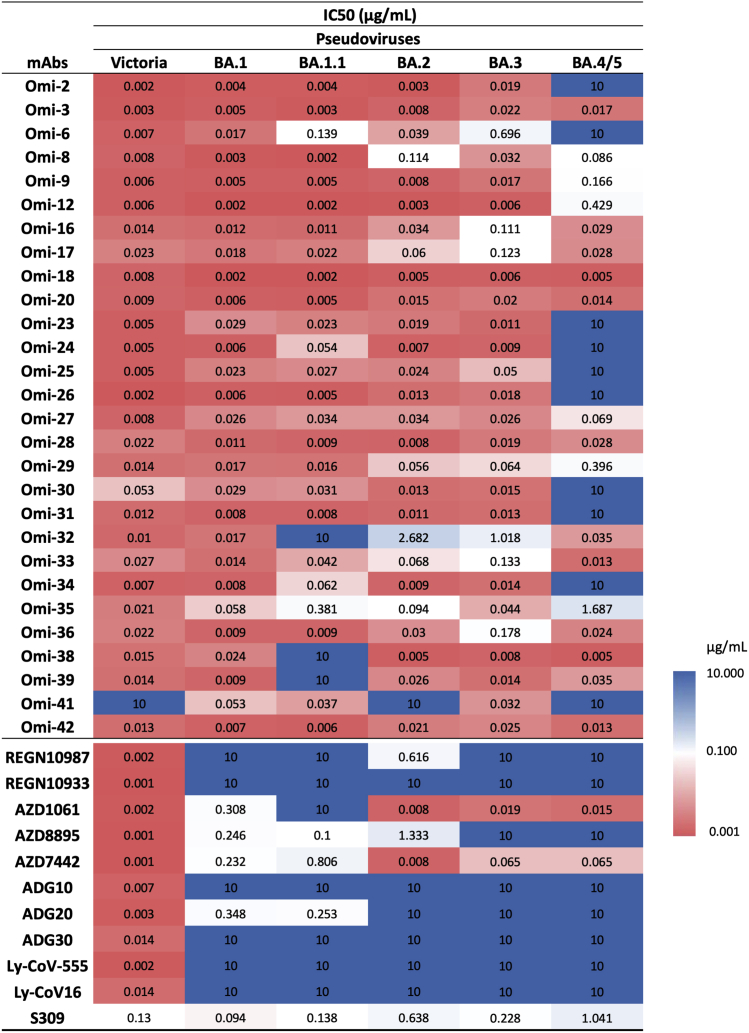
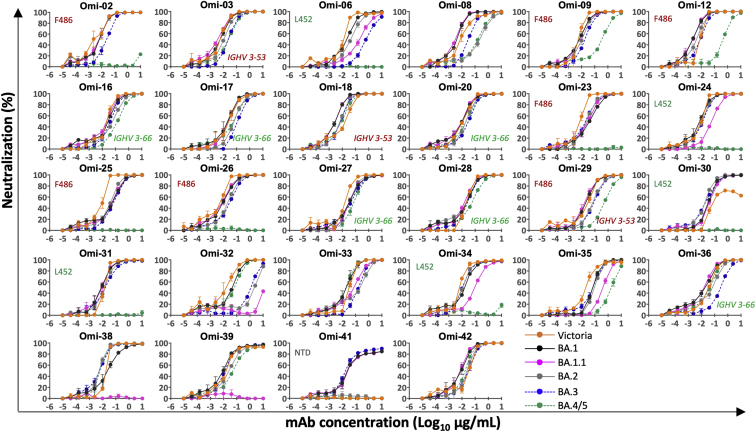
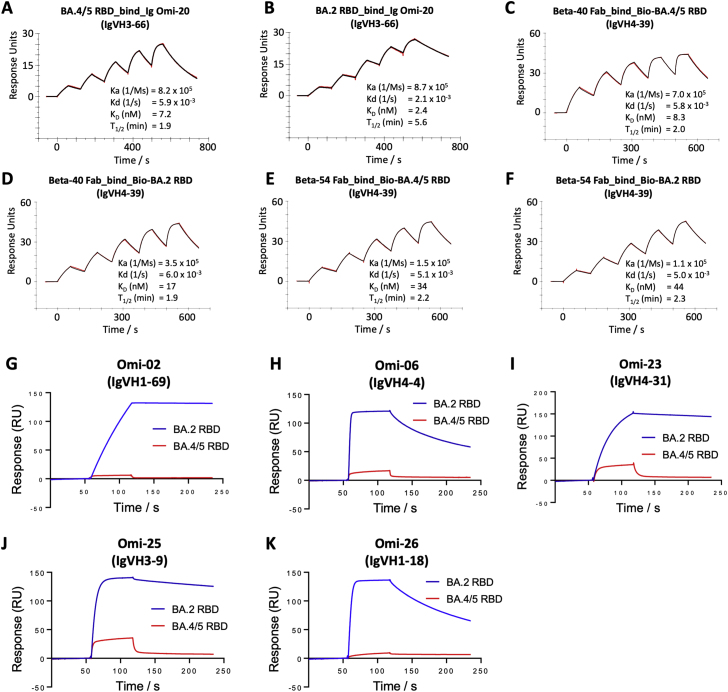
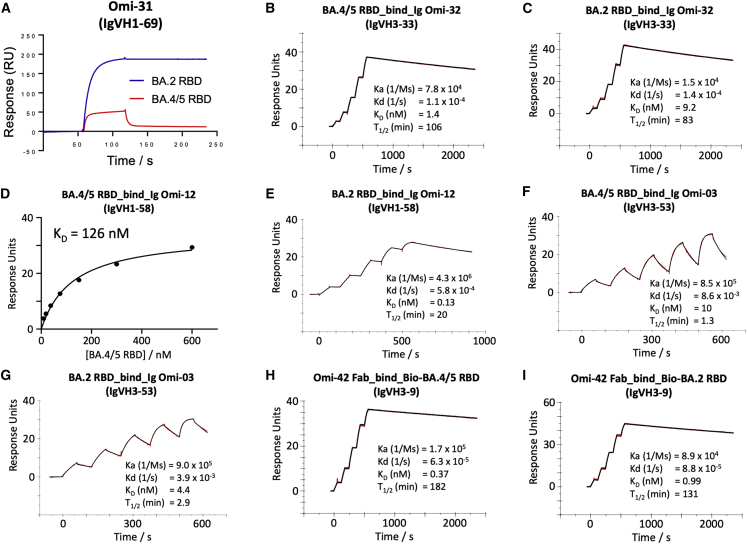
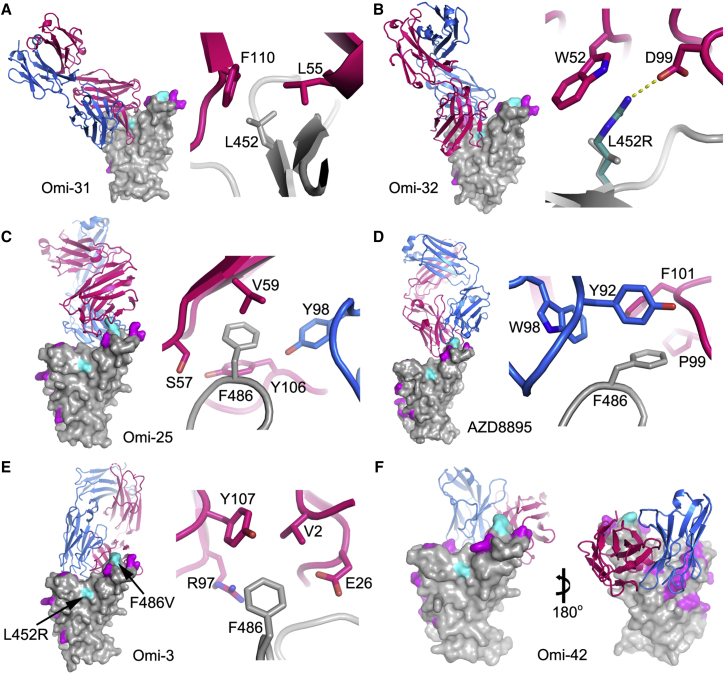
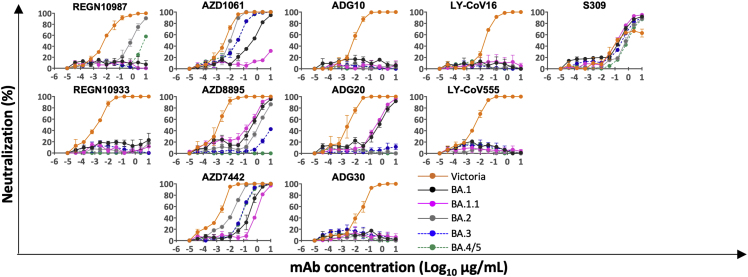
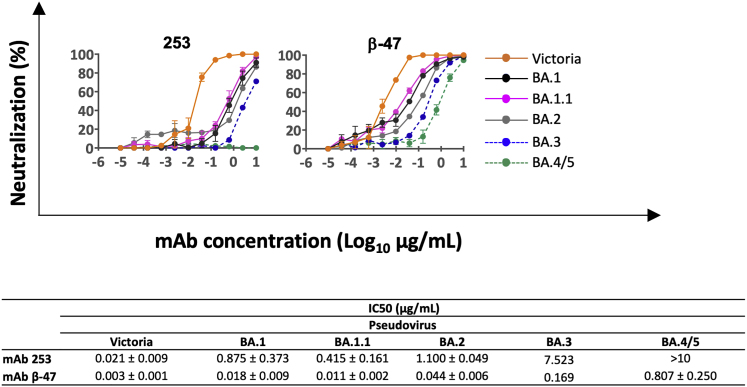
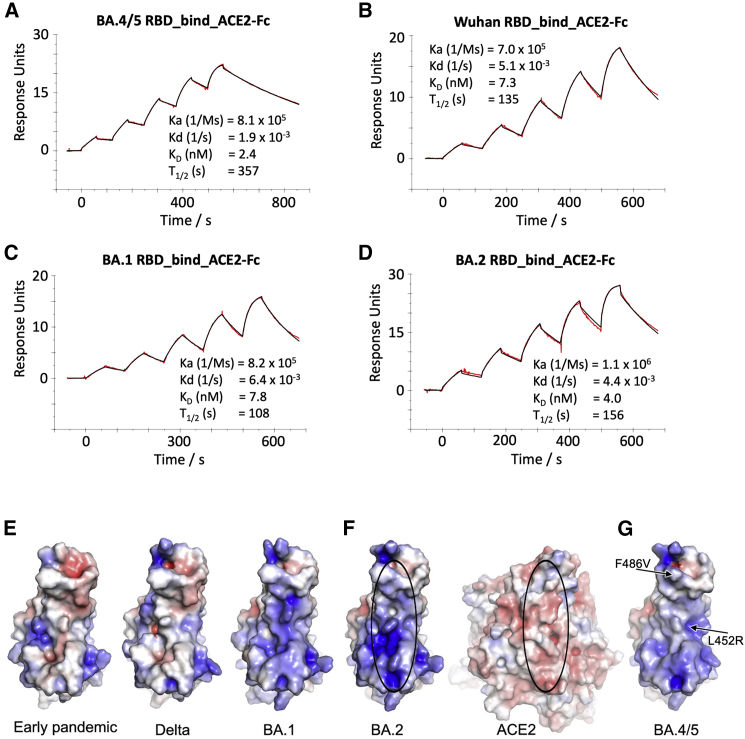

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
