

GTSF1 accelerates target RNA cleavage by PIWI-clade Argonaute proteins
- PMID: 35772669
- PMCID: PMC9385479
- DOI: 10.1038/s41586-022-05009-0
GTSF1 accelerates target RNA cleavage by PIWI-clade Argonaute proteins
Abstract
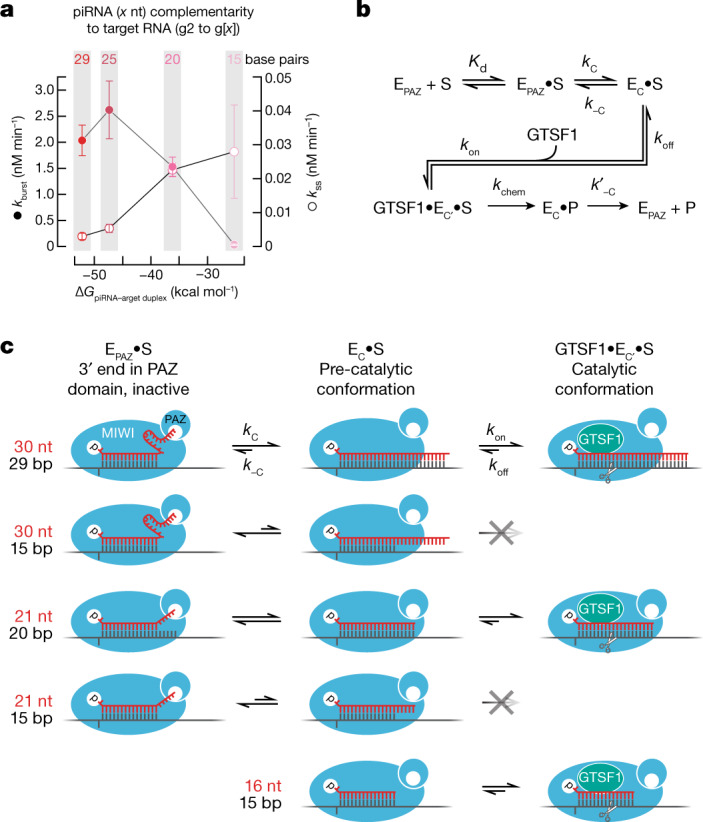
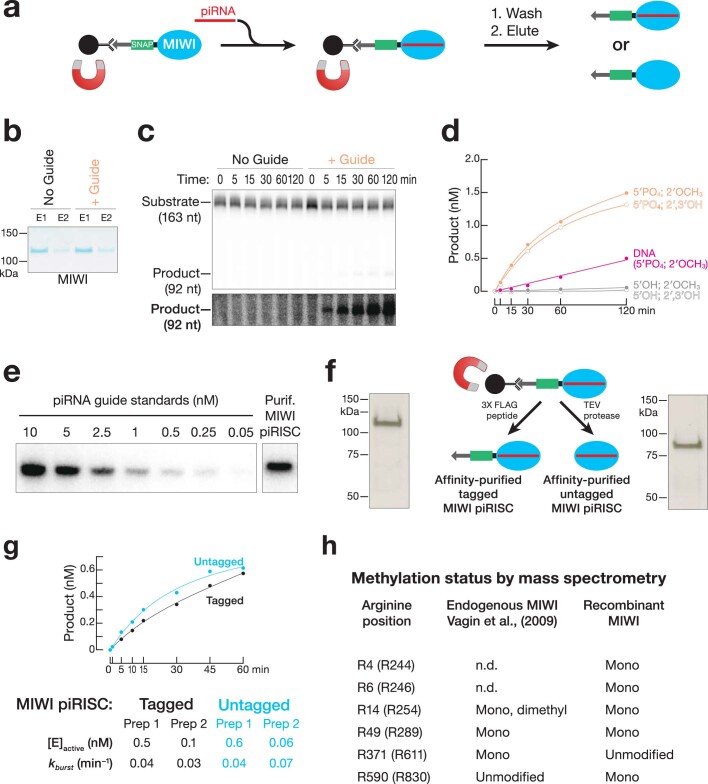
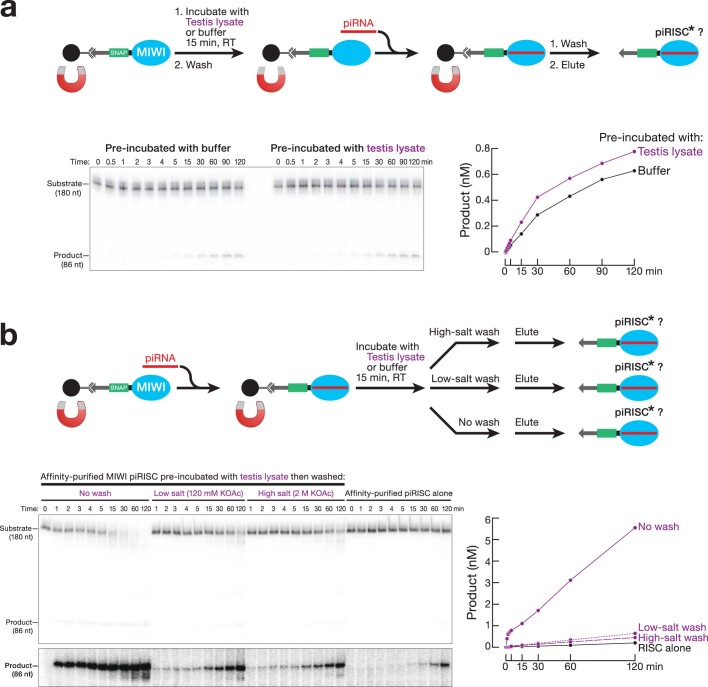
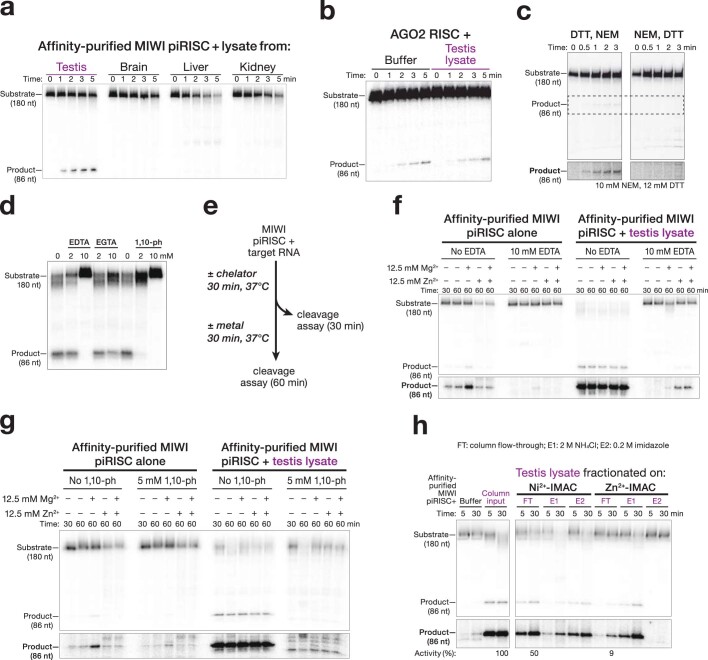
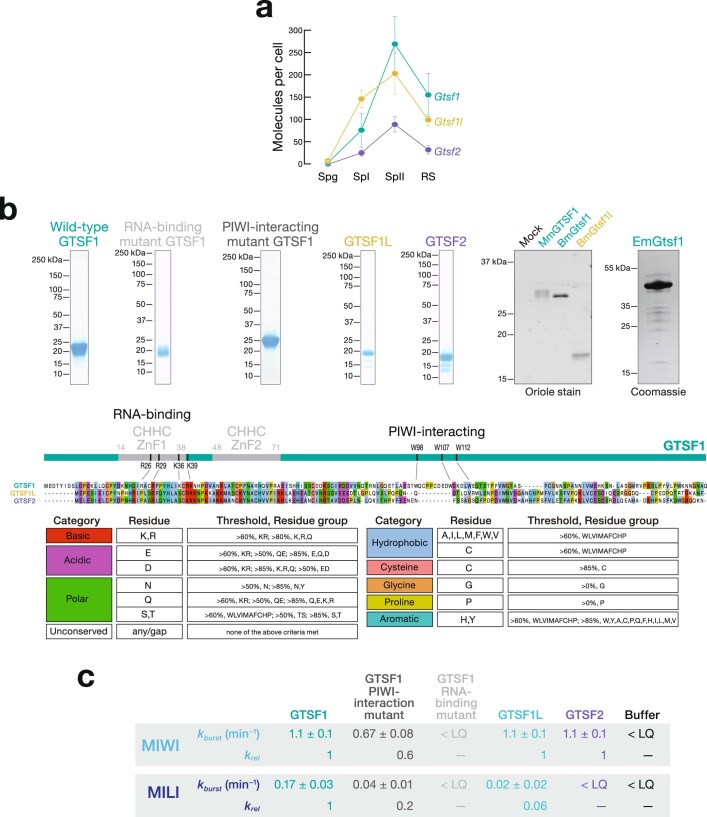
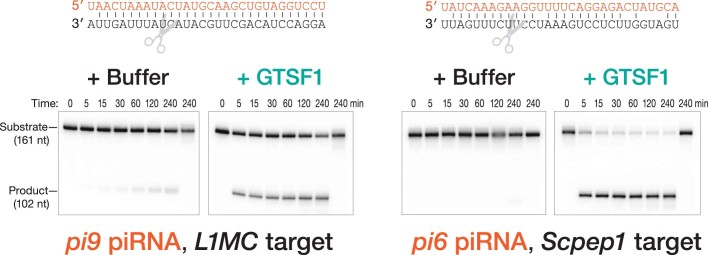
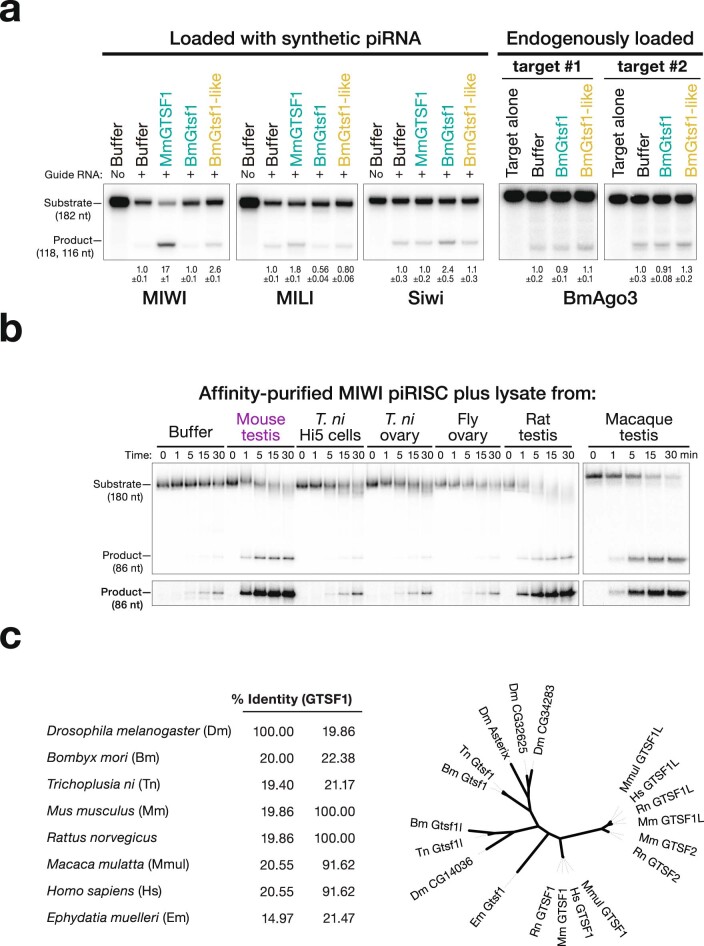
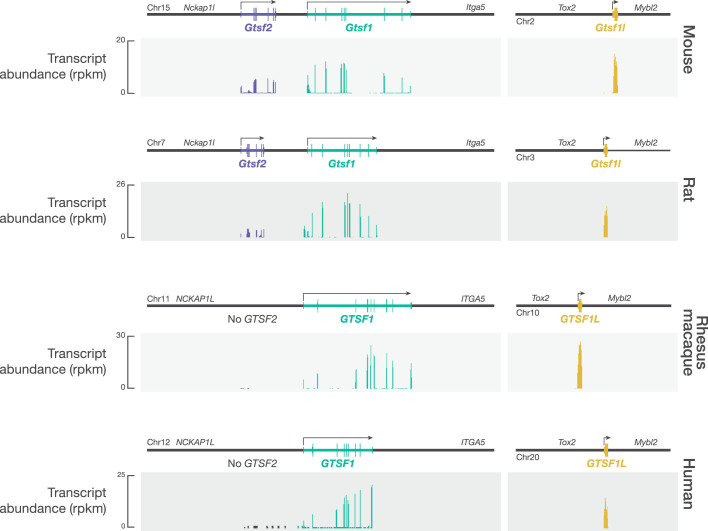
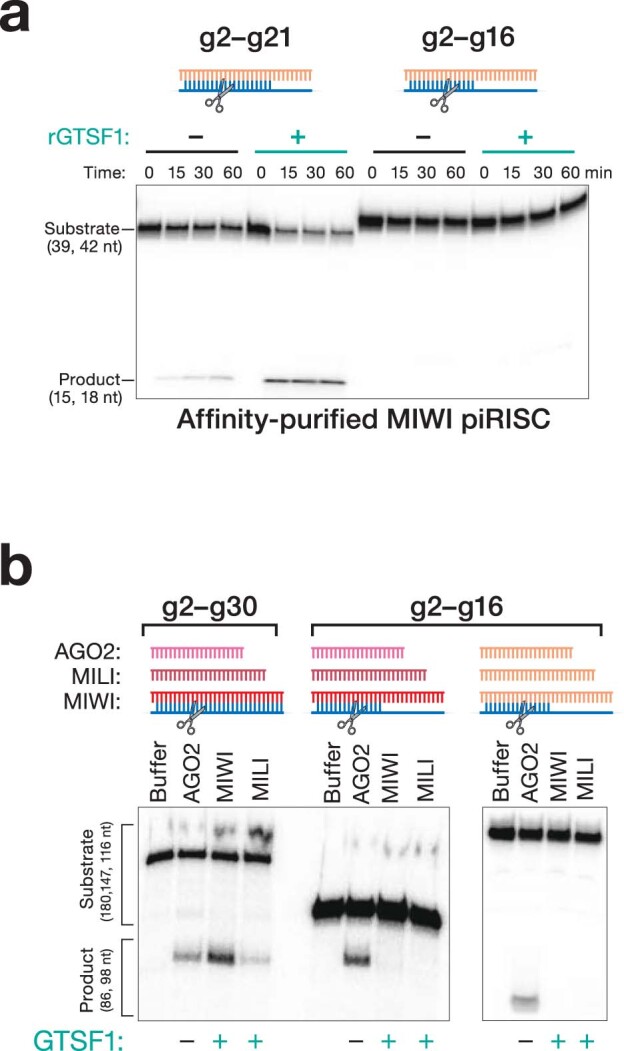
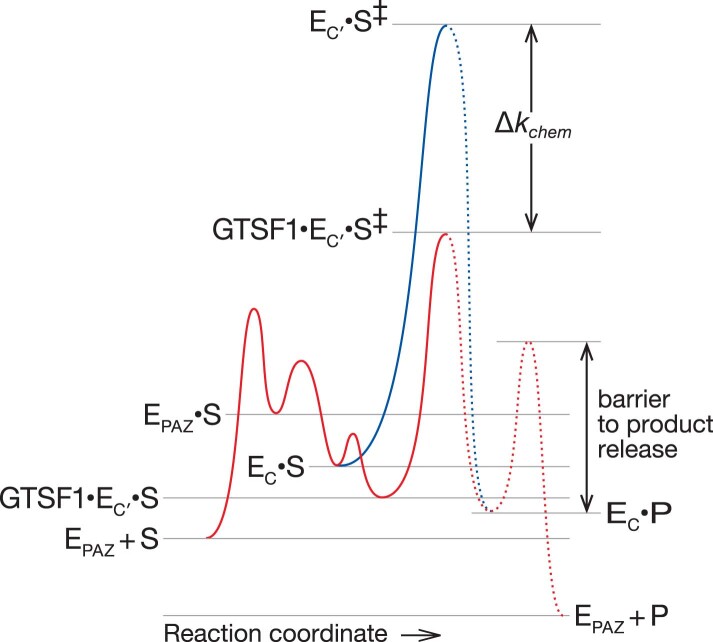
Argonaute proteins use nucleic acid guides to find and bind specific DNA or RNA target sequences. Argonaute proteins have diverse biological functions and many retain their ancestral endoribonuclease activity, cleaving the phosphodiester bond between target nucleotides t10 and t11. In animals, the PIWI proteins-a specialized class of Argonaute proteins-use 21-35 nucleotide PIWI-interacting RNAs (piRNAs) to direct transposon silencing, protect the germline genome, and regulate gene expression during gametogenesis1. The piRNA pathway is required for fertility in one or both sexes of nearly all animals. Both piRNA production and function require RNA cleavage catalysed by PIWI proteins. Spermatogenesis in mice and other placental mammals requires three distinct, developmentally regulated PIWI proteins: MIWI (PIWIL1), MILI (PIWIL2) and MIWI22-4 (PIWIL4). The piRNA-guided endoribonuclease activities of MIWI and MILI are essential for the production of functional sperm5,6. piRNA-directed silencing in mice and insects also requires GTSF1, a PIWI-associated protein of unknown function7-12. Here we report that GTSF1 potentiates the weak, intrinsic, piRNA-directed RNA cleavage activities of PIWI proteins, transforming them into efficient endoribonucleases. GTSF1 is thus an example of an auxiliary protein that potentiates the catalytic activity of an Argonaute protein.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
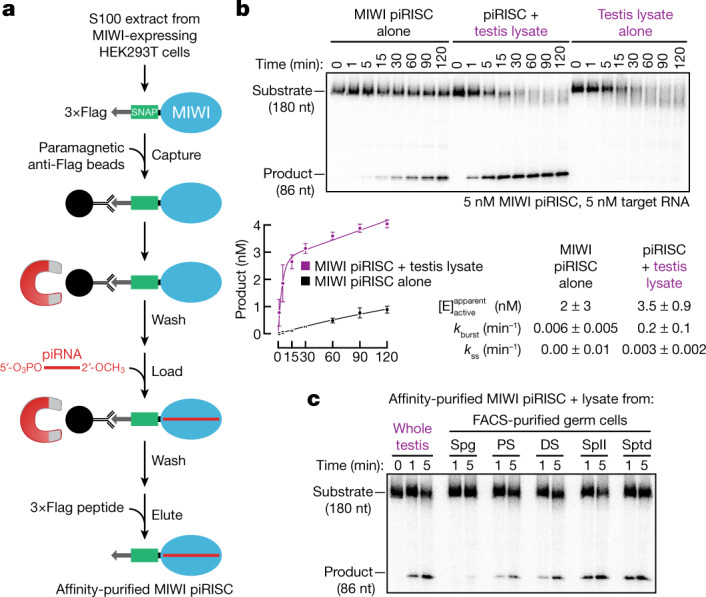
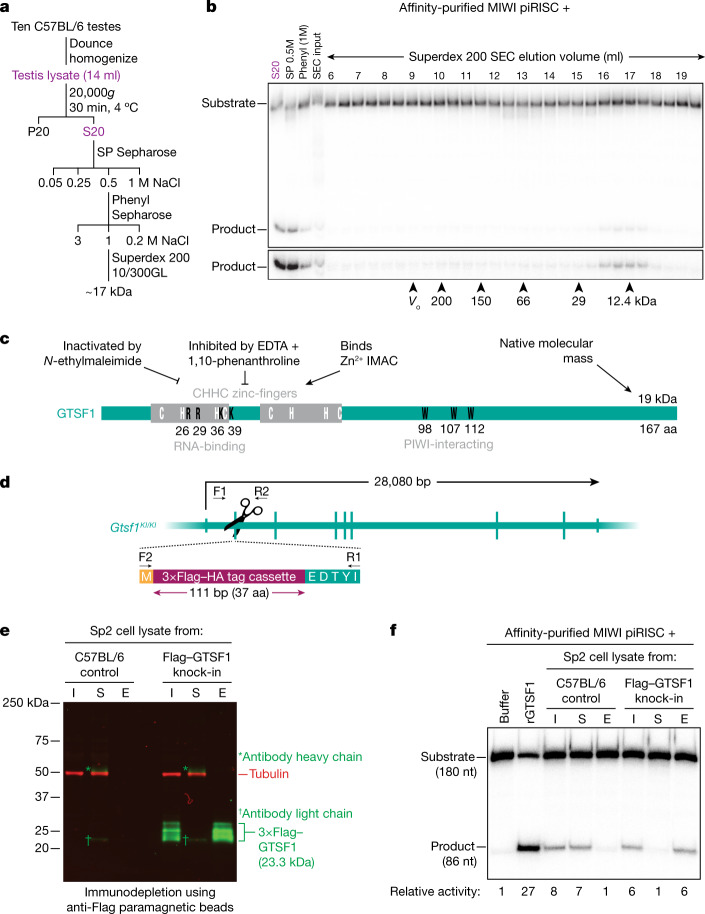
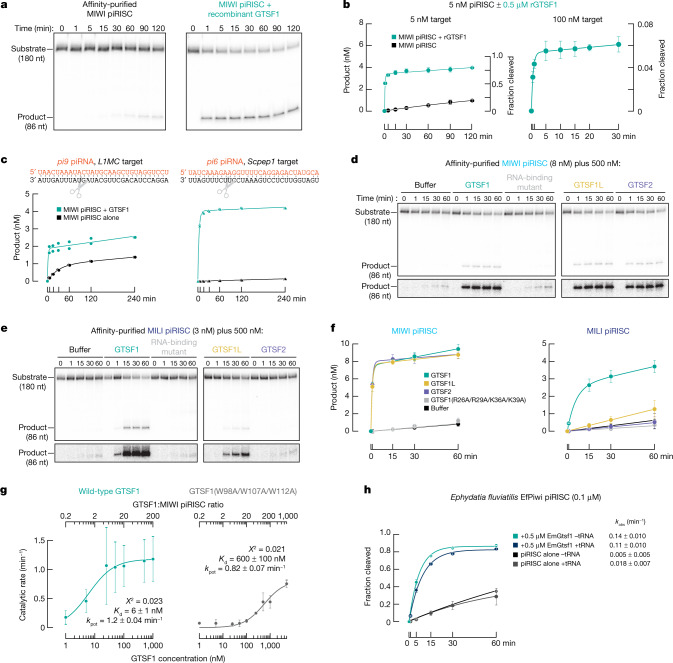
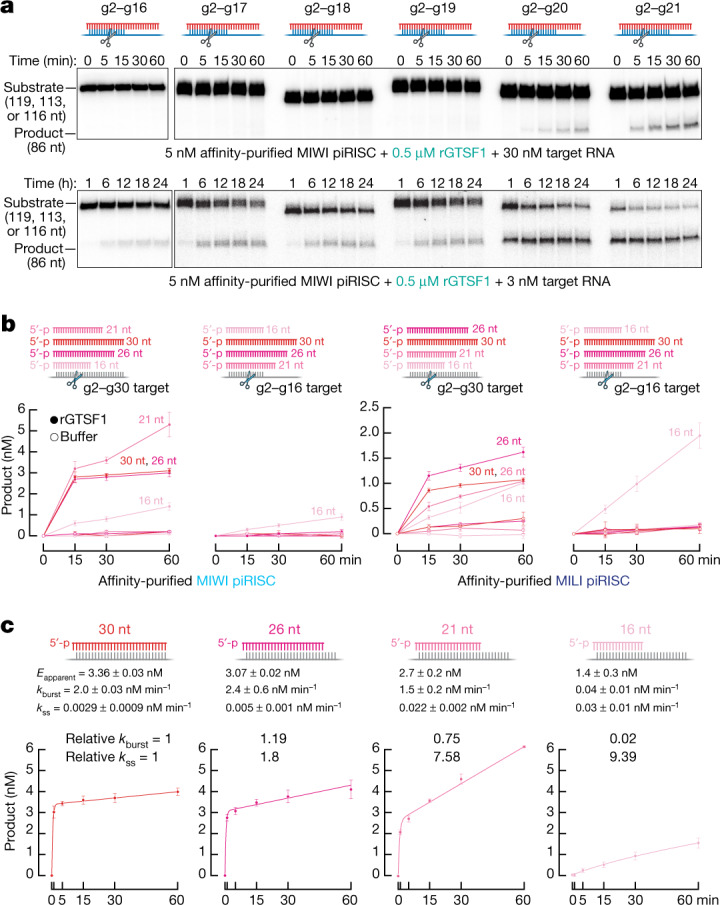
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
