

Cardiac glycoside-mediated turnover of Na, K-ATPases as a rational approach to reducing cell surface levels of the cellular prion protein
- PMID: 35776750
- PMCID: PMC9249225
- DOI: 10.1371/journal.pone.0270915
Cardiac glycoside-mediated turnover of Na, K-ATPases as a rational approach to reducing cell surface levels of the cellular prion protein
Abstract
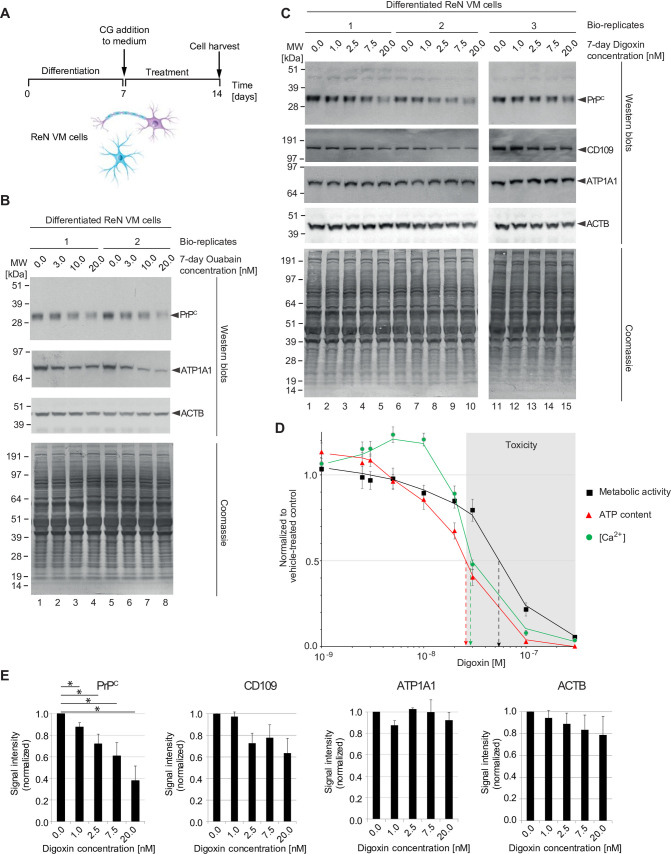
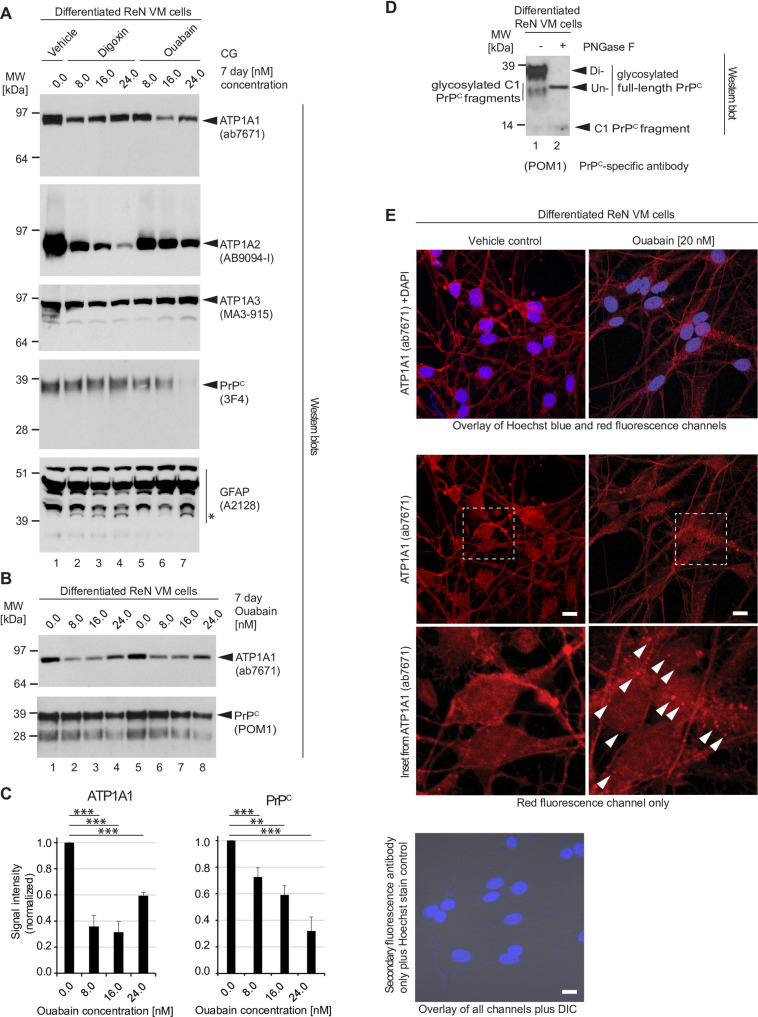
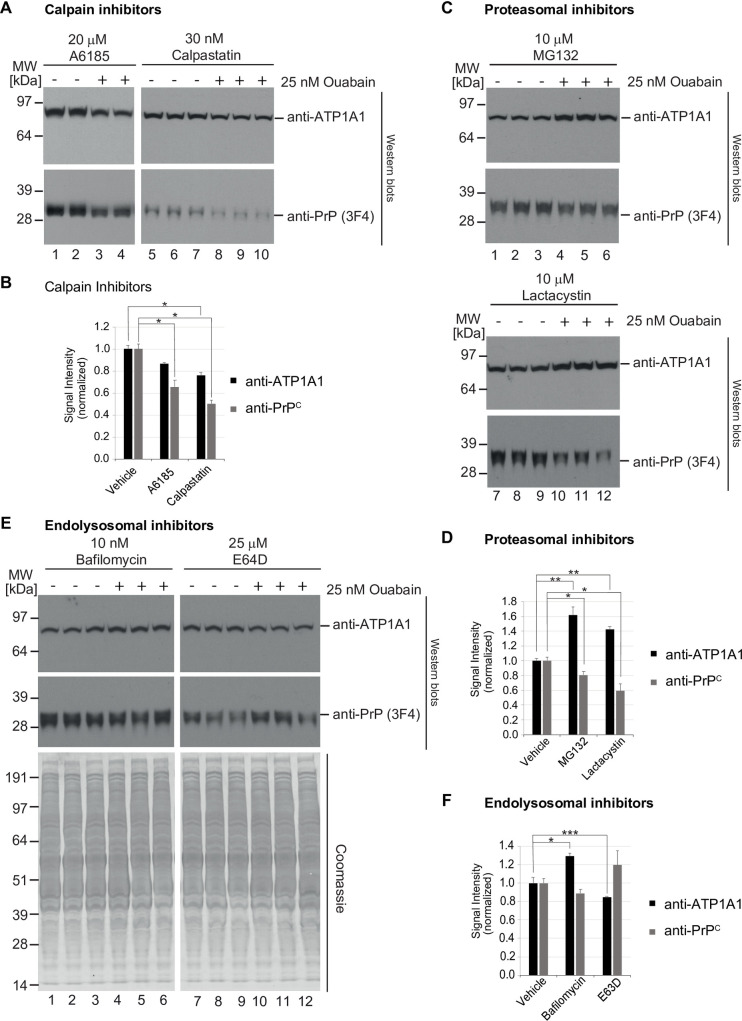
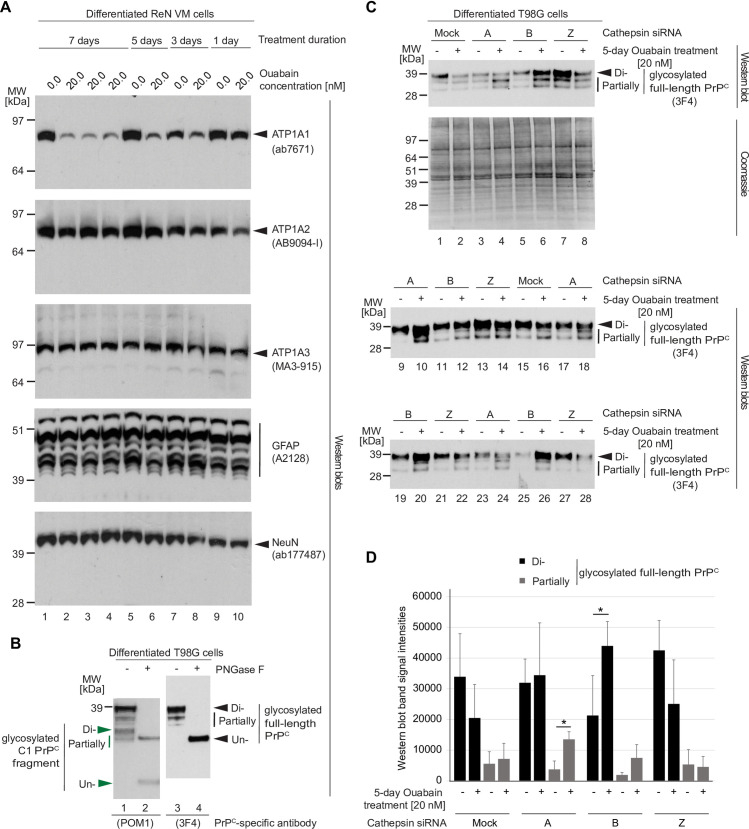
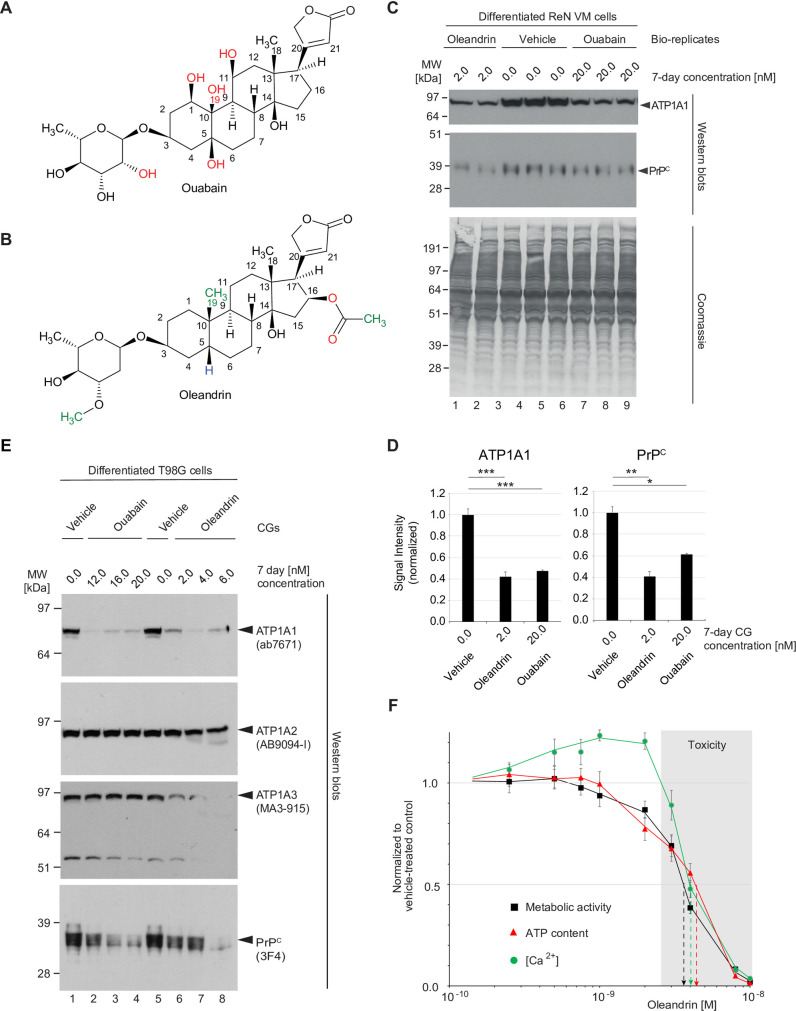
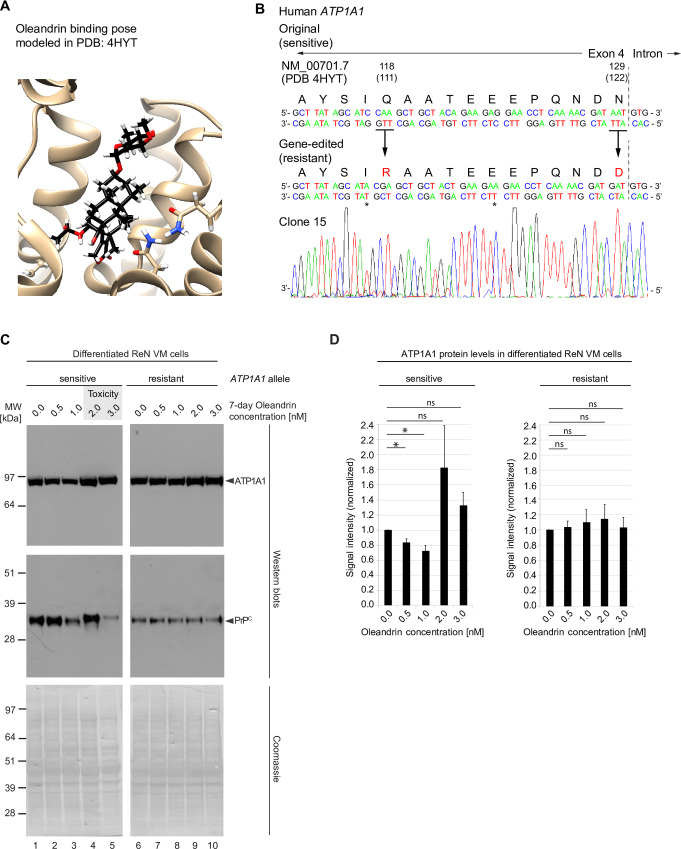
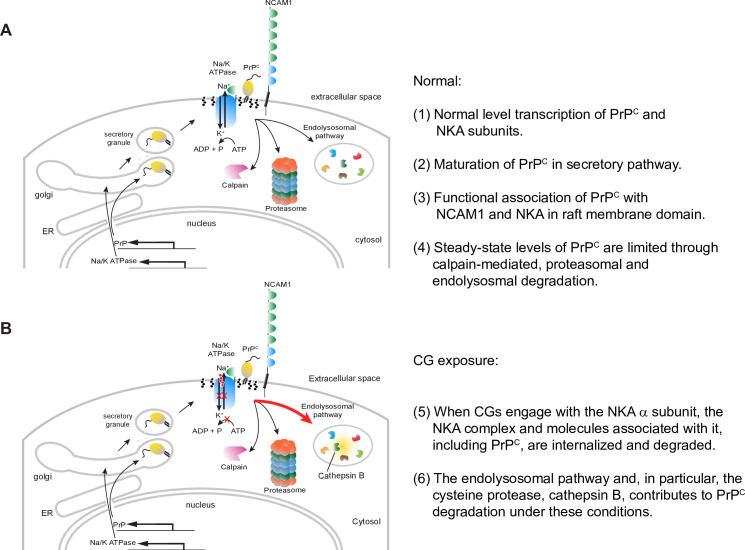
It is widely anticipated that a reduction of brain levels of the cellular prion protein (PrPC) can prolong survival in a group of neurodegenerative diseases known as prion diseases. To date, efforts to decrease steady-state PrPC levels by targeting this protein directly with small molecule drug-like compounds have largely been unsuccessful. Recently, we reported Na,K-ATPases to reside in immediate proximity to PrPC in the brain, unlocking an opportunity for an indirect PrPC targeting approach that capitalizes on the availability of potent cardiac glycosides (CGs). Here, we report that exposure of human co-cultures of neurons and astrocytes to non-toxic nanomolar levels of CGs causes profound reductions in PrPC levels. The mechanism of action underpinning this outcome relies primarily on a subset of CGs engaging the ATP1A1 isoform, one of three α subunits of Na,K-ATPases expressed in brain cells. Upon CG docking to ATP1A1, the ligand receptor complex, and PrPC along with it, is internalized by the cell. Subsequently, PrPC is channeled to the lysosomal compartment where it is digested in a manner that can be rescued by silencing the cysteine protease cathepsin B. These data signify that the repurposing of CGs may be beneficial for the treatment of prion disorders.
Conflict of interest statement
GS, MM and DW declare that they are co-inventors on a provisional patent (United States Provisional Application No. 63/159,289) named ‘Compounds and methods to treat prion and related diseases’ describing the use of cardiac glycosides for the purpose of reducing steady-state levels of PrPC. No other competing interests exist, and this provisional patent does not alter the adherence of these authors to all PLOS ONE policies on sharing data and materials.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
