

Evolutionary bioenergetics of ciliates
- PMID: 35778890
- PMCID: PMC11336482
- DOI: 10.1111/jeu.12934
Evolutionary bioenergetics of ciliates
Abstract
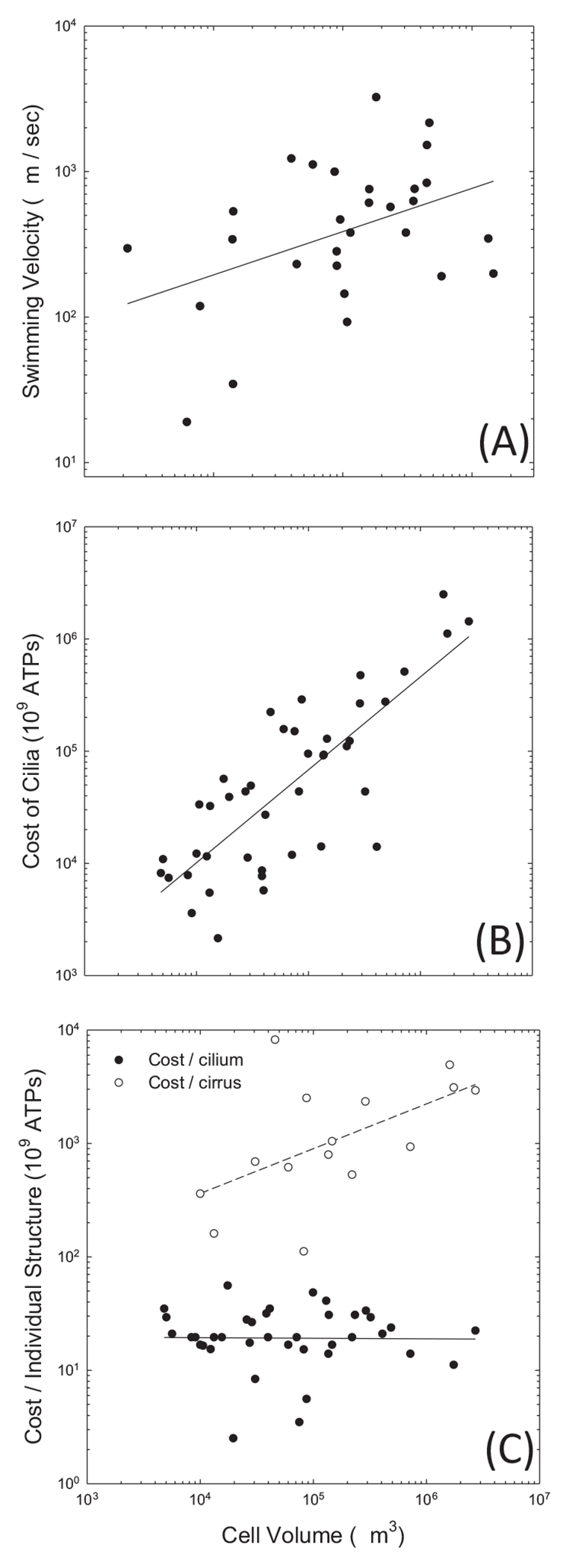
Understanding why various organisms evolve alternative ways of living requires information on both the fitness advantages of phenotypic modifications and the costs of constructing and operating cellular features. Although the former has been the subject of a myriad of ecological studies, almost no attention has been given to how organisms allocate resources to alternative structures and functions. We address these matters by capitalizing on an array of observations on diverse ciliate species and from the emerging field of evolutionary bioenergetics. A relatively robust and general estimator for the total cost of a cell per cell cycle (in units of ATP equivalents) is provided, and this is then used to understand how the magnitudes of various investments scale with cell size. Among other things, we examine the costs associated with the large macronuclear genomes of ciliates, as well as ribosomes, various internal membranes, osmoregulation, cilia, and swimming activities. Although a number of uncertainties remain, the general approach taken may serve as blueprint for expanding this line of work to additional traits and phylogenetic lineages.
Keywords: Paramecium; Tetrahymena; bioenergetics; ciliates; evolutionary cell biology; osmoregulation; ribosomes; swimming motility.
© 2022 International Society of Protistologists.
Figures


References
-
- Allen RD, Schroeder CC & Fok AK (1992) Endosomal system of Paramecium: coated pits to early endosomes. Journal of Cell Science, 101, 449–461. - PubMed
-
- Barsanti L, Coltelli P, Evangelista V, Frassanito AM & Gualtieri P (2016) Swimming patterns of the quadriflagellate Tetraflagellochloris mauritanica (Chlamy- domonadales, Chlorophyceae). Journal of Phycology, 52, 209–218. - PubMed
-
- Bauchop T & Elsden SR (1960) The growth of micro-organisms in relation to their energy supply. Journal of General Microbiology, 23, 457–469. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
