

Overexpression of MRX9 impairs processing of RNAs encoding mitochondrial oxidative phosphorylation factors COB and COX1 in yeast
- PMID: 35779633
- PMCID: PMC9307953
- DOI: 10.1016/j.jbc.2022.102214
Overexpression of MRX9 impairs processing of RNAs encoding mitochondrial oxidative phosphorylation factors COB and COX1 in yeast
Abstract
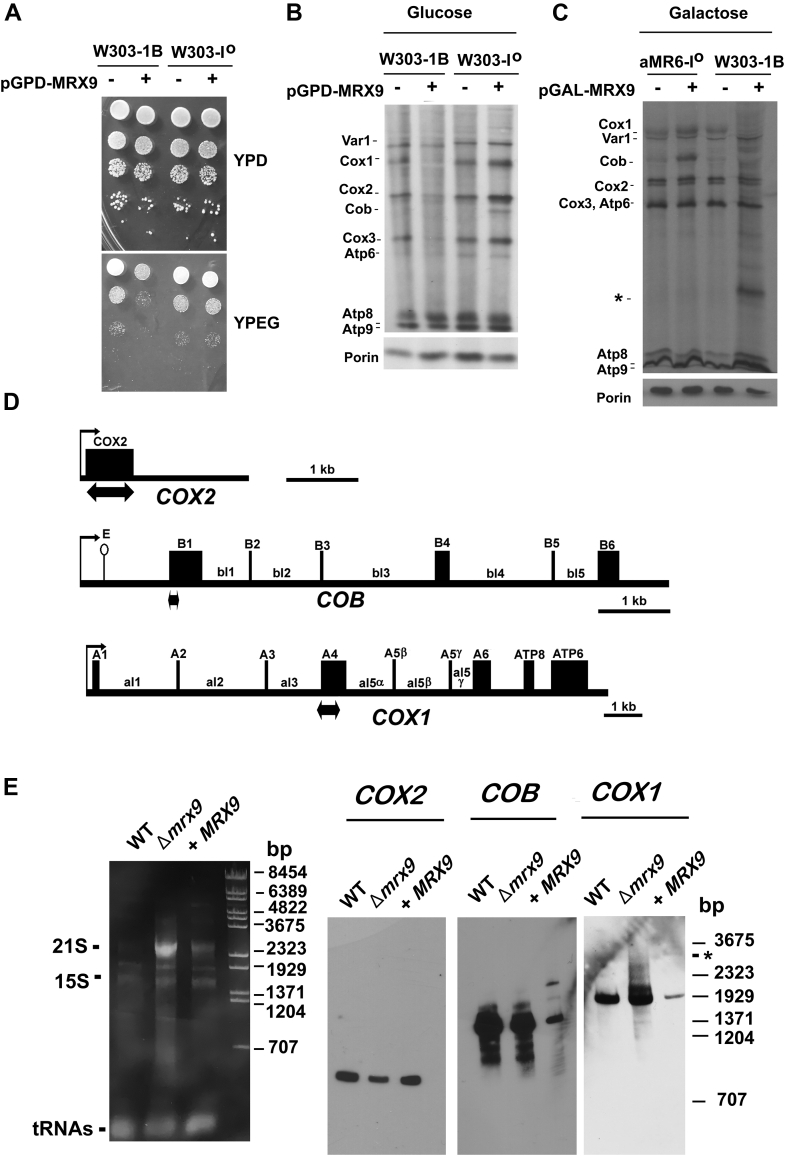
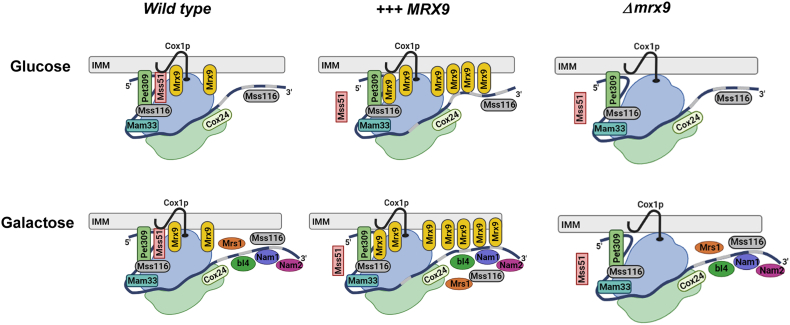
Mitochondrial translation is a highly regulated process, and newly synthesized mitochondrial products must first associate with several nuclear-encoded auxiliary factors to form oxidative phosphorylation complexes. The output of mitochondrial products should therefore be in stoichiometric equilibrium with the nuclear-encoded products to prevent unnecessary energy expense or the accumulation of pro-oxidant assembly modules. In the mitochondrial DNA of Saccharomyces cerevisiae, COX1 encodes subunit 1 of the cytochrome c oxidase and COB the central core of the cytochrome bc1 electron transfer complex; however, factors regulating the expression of these mitochondrial products are not completely described. Here, we identified Mrx9p as a new factor that controls COX1 and COB expression. We isolated MRX9 in a screen for mitochondrial factors that cause poor accumulation of newly synthesized Cox1p and compromised transition to the respiratory metabolism. Northern analyses indicated lower levels of COX1 and COB mature mRNAs accompanied by an accumulation of unprocessed transcripts in the presence of excess Mrx9p. In a strain devoid of mitochondrial introns, MRX9 overexpression did not affect COX1 and COB translation or respiratory adaptation, implying Mrx9p regulates processing of COX1 and COB RNAs. In addition, we found Mrx9p was localized in the mitochondrial inner membrane, facing the matrix, as a portion of it cosedimented with mitoribosome subunits and its removal or overexpression altered Mss51p sedimentation. Finally, we showed accumulation of newly synthesized Cox1p in the absence of Mrx9p was diminished in cox14 null mutants. Taken together, these data indicate a regulatory role of Mrx9p in COX1 RNA processing.
Keywords: Saccharomyces cerevisiae; intron processing; mitochondrial translation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures







References
-
- Ott M., Amunts A., Brown A. Organization and regulation of mitochondrial protein synthesis. Annu. Rev. Biochem. 2016;85:77–101. - PubMed
-
- Dujon B. Mitochondrial genetics revisited. Yeast. 2020;37:191–205. - PubMed
-
- Derbikova K.S., Levitsky S.A., Chicherin I.V., Vinogradova E.N., Kamenski P.A. Activation of yeast mitochondrial translation: who is in charge? Biochemistry. 2018;83:87–97. - PubMed
-
- Khalimonchuk O., Bird A., Winge D.R. Evidence for a pro-oxidant intermediate in the assembly of cytochrome oxidase. J. Biol. Chem. 2007;282:17442–17449. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
