

Pathological characterization of a novel mouse model expressing the PD-linked CHCHD2-T61I mutation
- PMID: 35786718
- PMCID: PMC9703812
- DOI: 10.1093/hmg/ddac083
Pathological characterization of a novel mouse model expressing the PD-linked CHCHD2-T61I mutation
Abstract
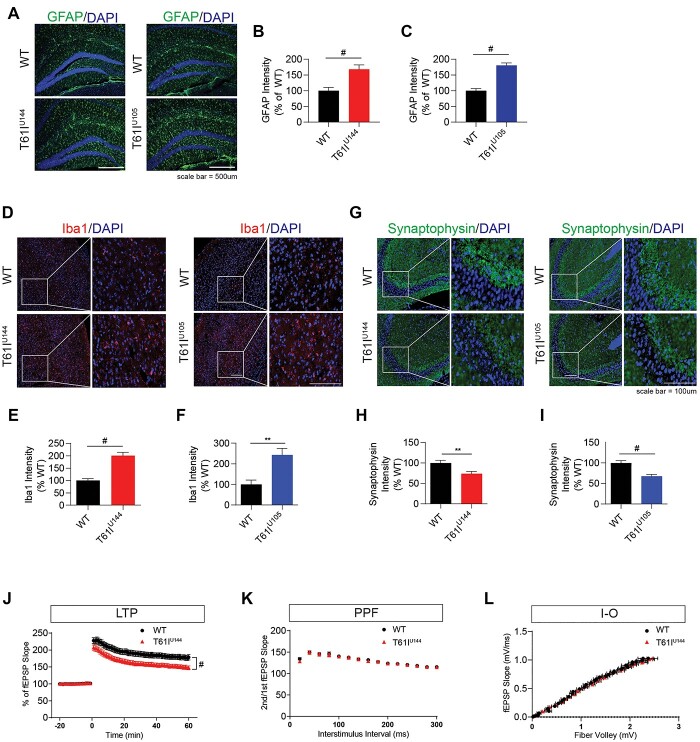
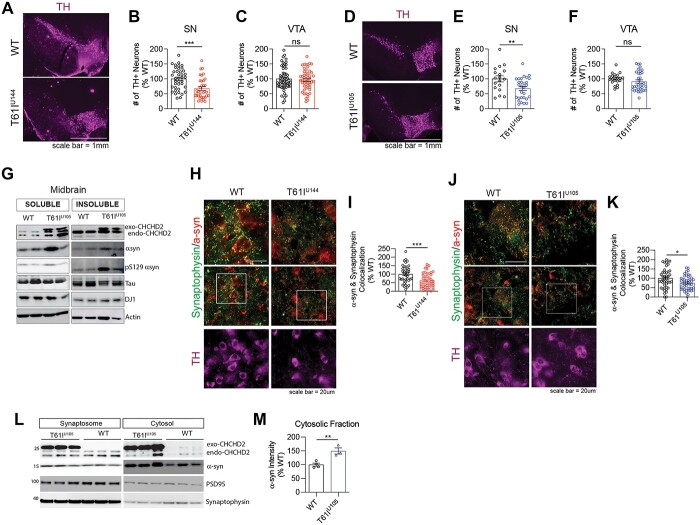
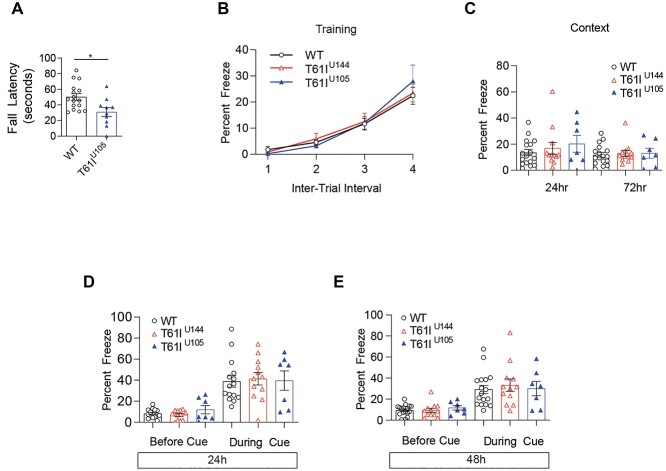
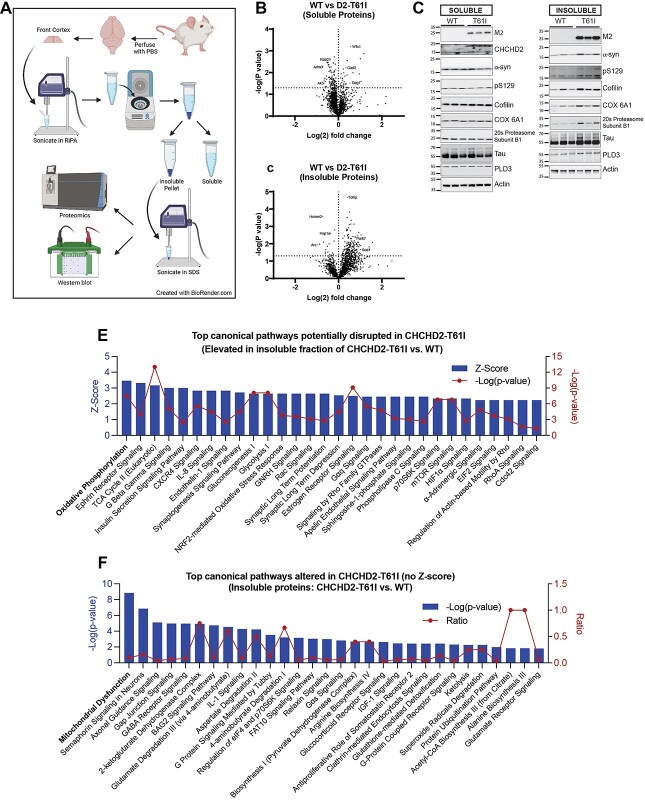
Coiled-coil-helix-coiled-coil-helix domain containing 2 (CHCHD2) is a mitochondrial protein that plays important roles in cristae structure, oxidative phosphorylation and apoptosis. Multiple mutations in CHCHD2 have been associated with Lewy body disorders (LBDs), such as Parkinson's disease (PD) and dementia with Lewy bodies, with the CHCHD2-T61I mutation being the most widely studied. However, at present, only CHCHD2 knockout or CHCHD2/CHCHD10 double knockout mouse models have been investigated. They do not recapitulate the pathology seen in patients with CHCHD2 mutations. We generated the first transgenic mouse model expressing the human PD-linked CHCHD2-T61I mutation driven by the mPrP promoter. We show that CHCHD2-T61I Tg mice exhibit perinuclear mitochondrial aggregates, neuroinflammation, and have impaired long-term synaptic plasticity associated with synaptic dysfunction. Dopaminergic neurodegeneration, a hallmark of PD, is also observed along with α-synuclein pathology. Significant motor dysfunction is seen with no changes in learning and memory at 1 year of age. A minor proportion of the CHCHD2-T61I Tg mice (~10%) show a severe motor phenotype consistent with human Pisa Syndrome, an atypical PD phenotype. Unbiased proteomics analysis reveals surprising increases in many insoluble proteins predominantly originating from mitochondria and perturbing multiple canonical biological pathways as assessed by ingenuity pathway analysis, including neurodegenerative disease-associated proteins such as tau, cofilin, SOD1 and DJ-1. Overall, CHCHD2-T61I Tg mice exhibit pathological and motor changes associated with LBDs, indicating that this model successfully captures phenotypes seen in human LBD patients with CHCHD2 mutations and demonstrates changes in neurodegenerative disease-associated proteins, which delineates relevant pathological pathways for further investigation.
© The Author(s) 2022. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
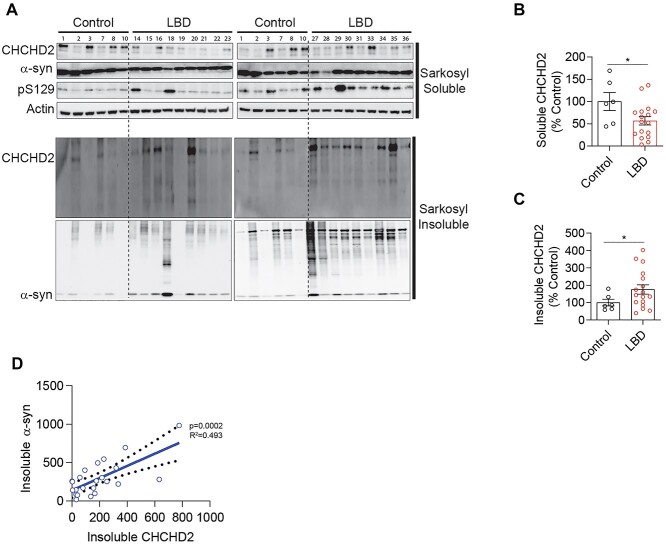
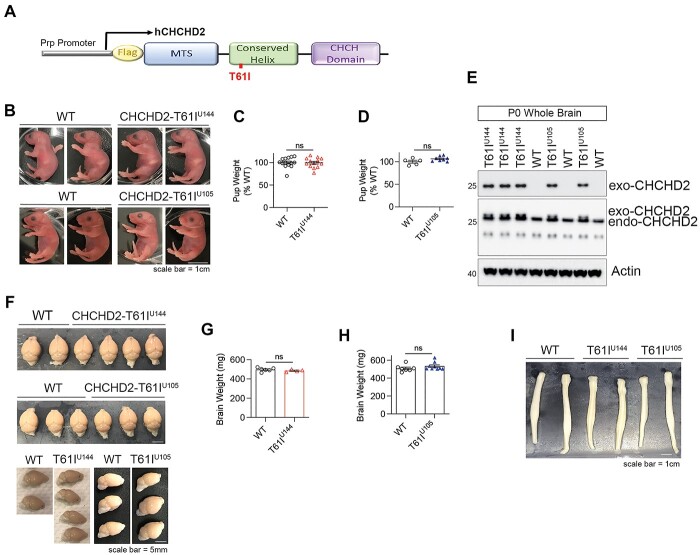
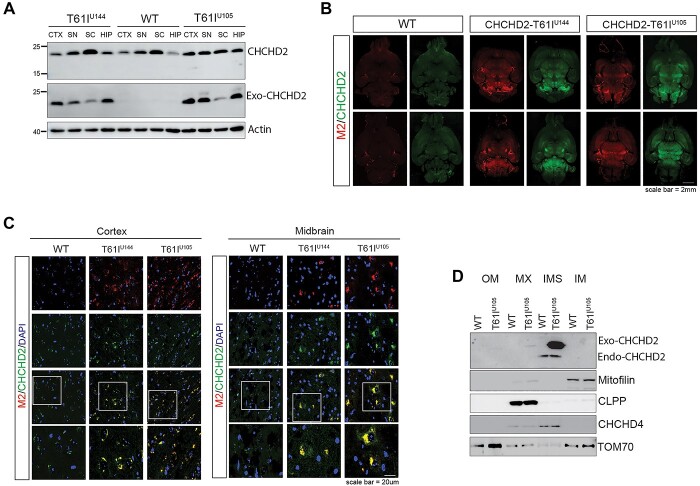
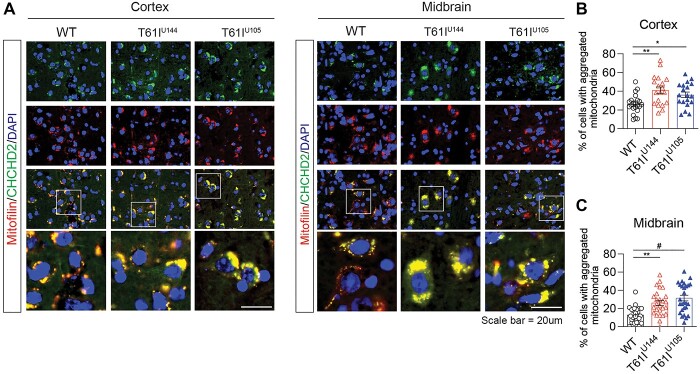
References
-
- Jellinger, K.A. (2018) Dementia with Lewy bodies and Parkinson's disease-dementia: current concepts and controversies. J. Neural Transm. (Vienna), 125, 615–650. - PubMed
-
- Spillantini, M.G., Schmidt, M.L., Lee, V.M., Trojanowski, J.Q., Jakes, R. and Goedert, M. (1997) Alpha-synuclein in Lewy bodies. Nature, 388, 839–840. - PubMed
-
- Beal, M.F. (1998) Mitochondrial dysfunction in neurodegenerative diseases. Biochim. Biophys. Acta, 1366, 211–223. - PubMed
-
- Lin, M.T. and Beal, M.F. (2006) Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature, 443, 787–795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
