

Transmission and diversity of Schistosoma haematobium and S. bovis and their freshwater intermediate snail hosts Bulinus globosus and B. nasutus in the Zanzibar Archipelago, United Republic of Tanzania
- PMID: 35788199
- PMCID: PMC9286283
- DOI: 10.1371/journal.pntd.0010585
Transmission and diversity of Schistosoma haematobium and S. bovis and their freshwater intermediate snail hosts Bulinus globosus and B. nasutus in the Zanzibar Archipelago, United Republic of Tanzania
Abstract
Background: The Zanzibar Archipelago (Pemba and Unguja islands) is targeted for the elimination of human urogenital schistosomiasis caused by infection with Schistosoma haematobium where the intermediate snail host is Bulinus globosus. Following multiple studies, it has remained unclear if B. nasutus (a snail species that occupies geographically distinct regions on the Archipelago) is involved in S. haematobium transmission on Zanzibar. Additionally, S. haematobium was thought to be the only Schistosoma species present on the Zanzibar Archipelago until the sympatric transmission of S. bovis, a parasite of ruminants, was recently identified. Here we re-assess the epidemiology of schistosomiasis on Pemba and Unguja together with the role and genetic diversity of the Bulinus spp. involved in transmission.
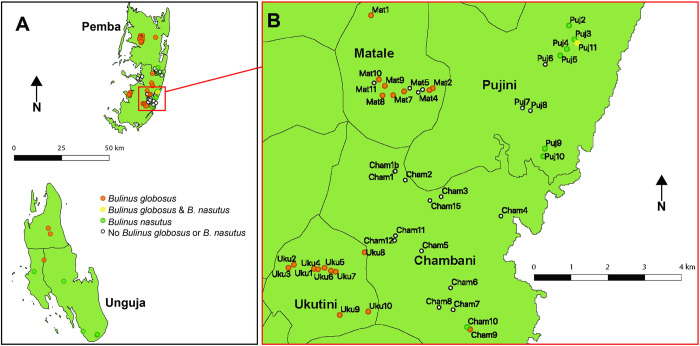
Methodology/principal findings: Malacological and parasitological surveys were conducted between 2016 and 2019. In total, 11,116 Bulinus spp. snails were collected from 65 of 112 freshwater bodies surveyed. Bulinus species identification were determined using mitochondrial cox1 sequences for a representative subset of collected Bulinus (n = 504) and together with archived museum specimens (n = 6), 433 B. globosus and 77 B. nasutus were identified. Phylogenetic analysis of cox1 haplotypes revealed three distinct populations of B. globosus, two with an overlapping distribution on Pemba and one on Unguja. For B. nasutus, only a single clade with matching haplotypes was observed across the islands and included reference sequences from Kenya. Schistosoma haematobium cercariae (n = 158) were identified from 12 infected B. globosus and one B. nasutus collected between 2016 and 2019 in Pemba, and cercariae originating from 69 Bulinus spp. archived in museum collections. Schistosoma bovis cercariae (n = 21) were identified from seven additional B. globosus collected between 2016 and 2019 in Pemba. By analysing a partial mitochondrial cox1 region and the nuclear ITS (1-5.8S-2) rDNA region of Schistosoma cercariae, we identified 18 S. haematobium and three S. bovis haplotypes representing populations associated with mainland Africa and the Indian Ocean Islands (Zanzibar, Madagascar, Mauritius and Mafia).
Conclusions/significance: The individual B. nasutus on Pemba infected with S. haematobium demonstrates that B. nasutus could also play a role in the local transmission of S. haematobium. We provide preliminary evidence that intraspecific variability of S. haematobium on Pemba may increase the transmission potential of S. haematobium locally due to the expanded intermediate host range, and that the presence of S. bovis complicates the environmental surveillance of schistosome infections.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- King CH, Kittur N, Binder S, Campbell CH, N’Goran EK, Meite A, et al.. Impact of Different Mass Drug Administration Strategies for Gaining and Sustaining Control of Schistosoma mansoni and Schistosoma haematobium Infection in Africa. Am J Trop Med Hyg. 2020;103: 14–23. doi: 10.4269/ajtmh.19-0829 - DOI - PMC - PubMed
-
- Allan F, Ame SM, Tian-Bi Y-NT, Hofkin B V, Webster BL, Diakité NR, et al.. Snail-Related Contributions from the Schistosomiasis Consortium for Operational Research and Evaluation Program Including Xenomonitoring, Focal Mollusciciding, Biological Control, and Modeling. Am J Trop Med Hyg. 2020. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
