

Cytarabine-induced differentiation of AML cells depends on Chk1 activation and shares the mechanism with inhibitors of DHODH and pyrimidine synthesis
- PMID: 35790845
- PMCID: PMC9256737
- DOI: 10.1038/s41598-022-15520-z
Cytarabine-induced differentiation of AML cells depends on Chk1 activation and shares the mechanism with inhibitors of DHODH and pyrimidine synthesis
Abstract
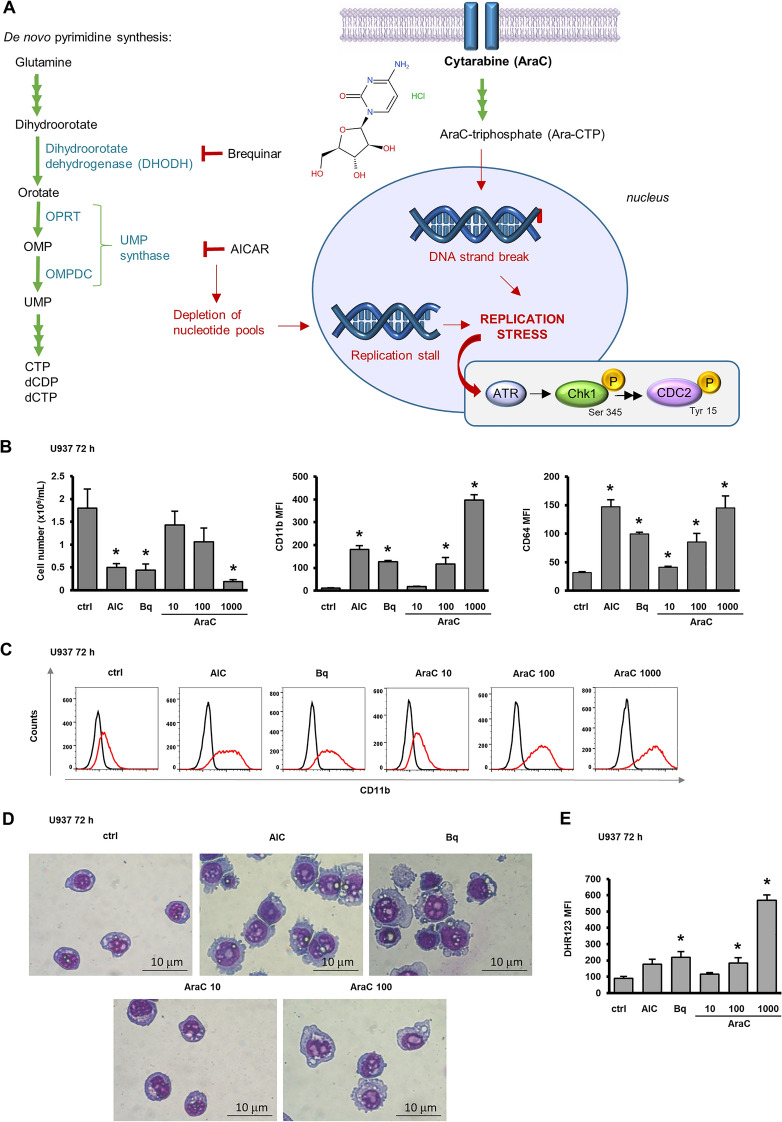
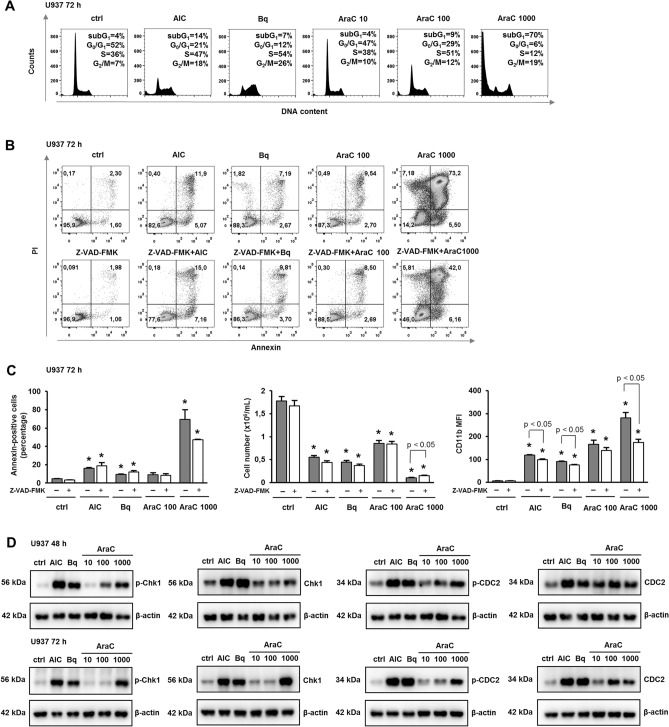
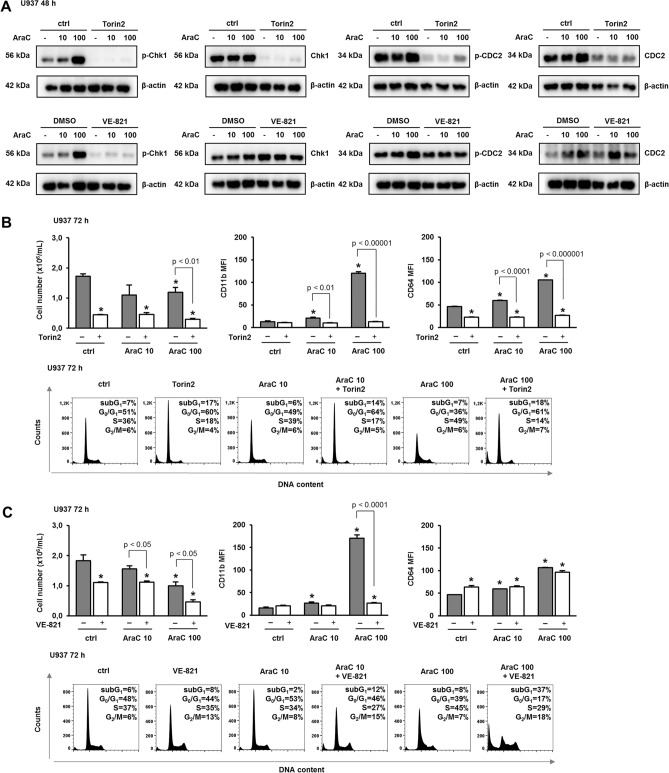
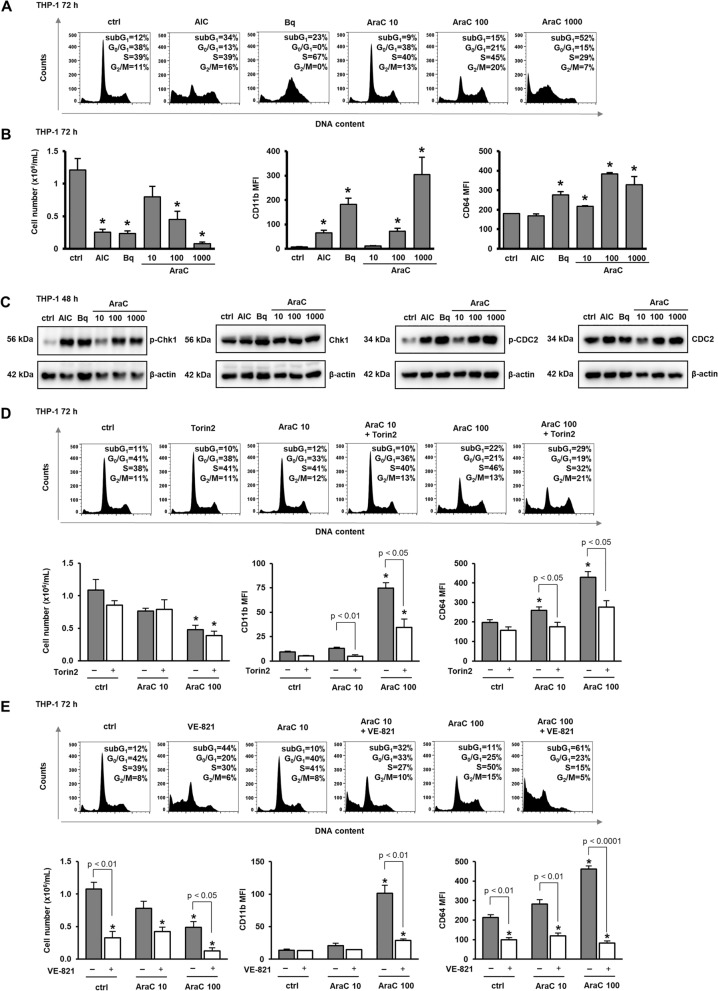
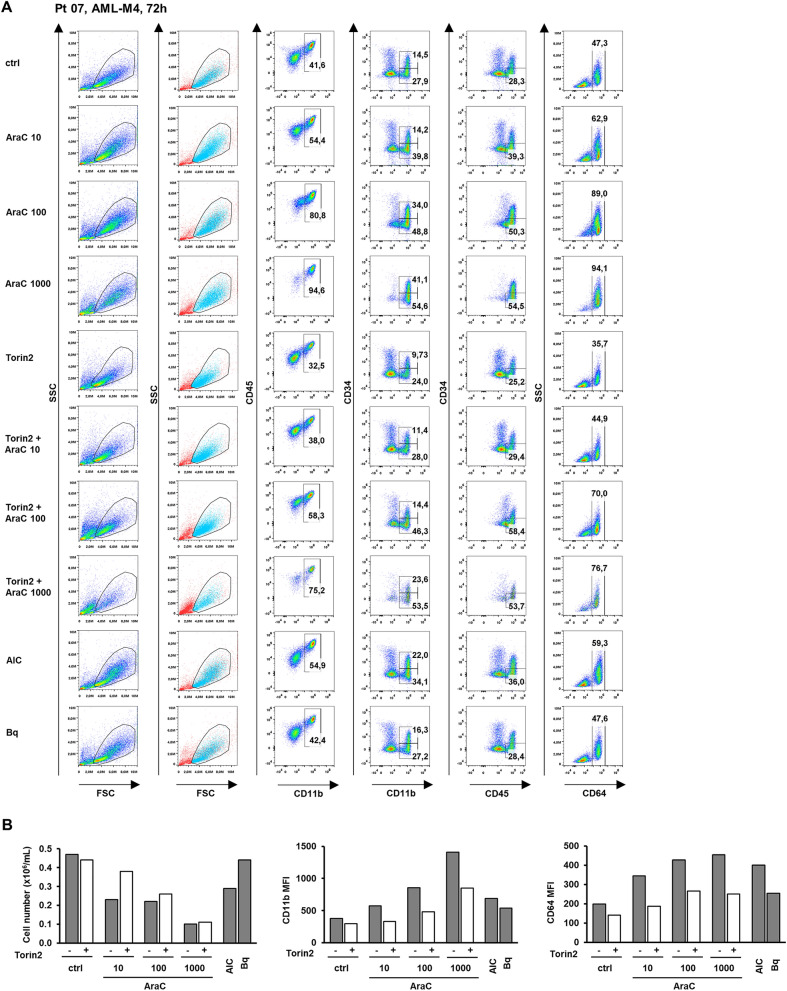
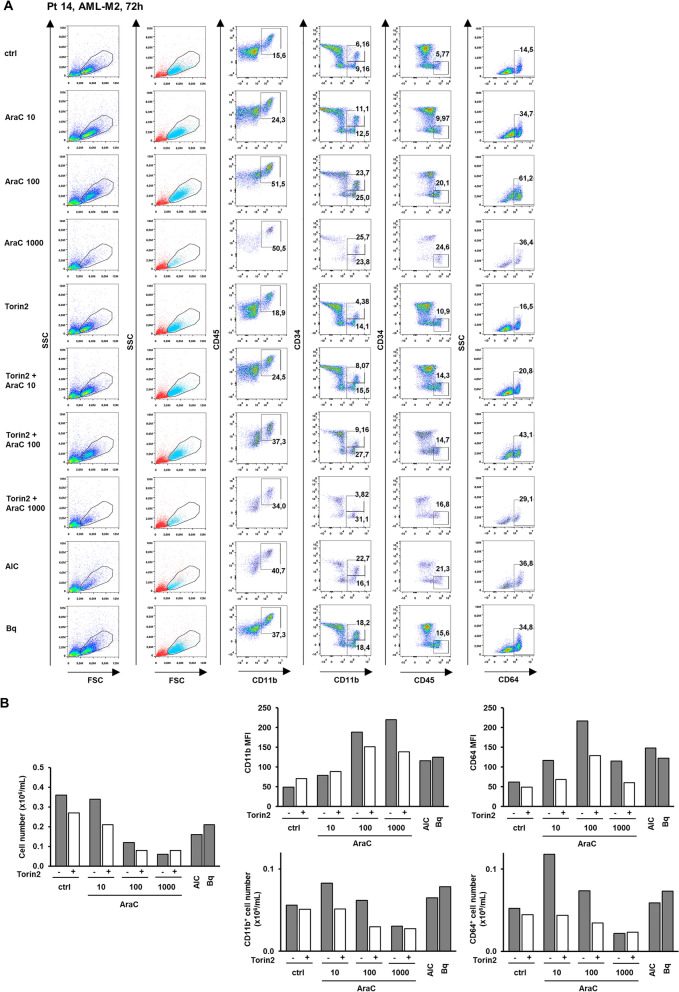
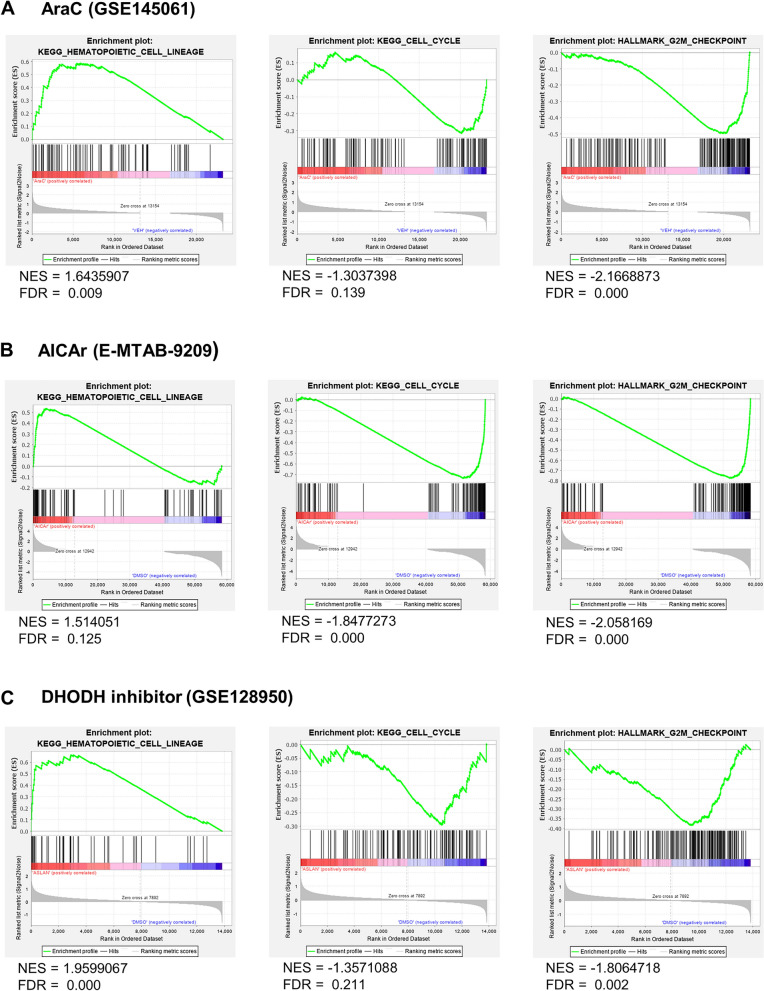
Acute myeloid leukemia (AML) is characterized by arrested differentiation making differentiation therapy a promising treatment strategy. Recent success of inhibitors of mutated isocitrate dehydrogenase (IDH) invigorated interest in differentiation therapy of AML so that several new drugs have been proposed, including inhibitors of dihydroorotate dehydrogenase (DHODH), an enzyme in pyrimidine synthesis. Cytarabine, a backbone of standard AML therapy, is known to induce differentiation at low doses, but the mechanism is not completely elucidated. We have previously reported that 5-aminoimidazole-4-carboxamide ribonucleoside (AICAr) and brequinar, a DHODH inhibitor, induced differentiation of myeloid leukemia by activating the ataxia telangiectasia and Rad3-related (ATR)/checkpoint kinase 1 (Chk1) via pyrimidine depletion. In this study, using immunoblotting, flow cytometry analyses, pharmacologic inhibitors and genetic inactivation of Chk1 in myeloid leukemia cell lines, we show that low dose cytarabine induces differentiation by activating Chk1. In addition, cytarabine induces differentiation ex vivo in a subset of primary AML samples that are sensitive to AICAr and DHODH inhibitor. The results of our study suggest that leukemic cell differentiation stimulated by low doses of cytarabine depends on the activation of Chk1 and thus shares the same pathway as pyrimidine synthesis inhibitors.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Short NJ, et al. Advances in the treatment of acute myeloid leukemia: New drugs and new challenges. Cancer Discov. 2020;10:506–525. doi: 10.1158/2159-8290.CD-19-1011. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
