

Neural regeneration therapy after spinal cord injury induces unique brain functional reorganizations in rhesus monkeys
- PMID: 35792748
- PMCID: PMC9272921
- DOI: 10.1080/07853890.2022.2089728
Neural regeneration therapy after spinal cord injury induces unique brain functional reorganizations in rhesus monkeys
Abstract
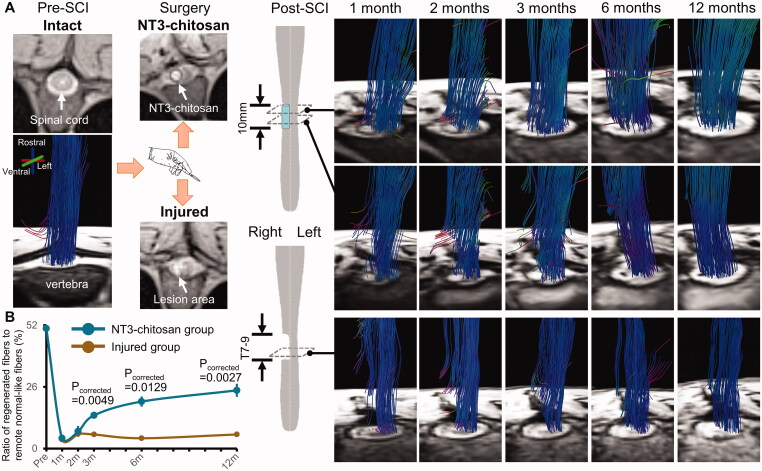
Purpose: Spinal cord injury (SCI) destroys the sensorimotor pathway and induces brain plasticity. However, the effect of treatment-induced spinal cord tissue regeneration on brain functional reorganization remains unclear. This study was designed to investigate the large-scale functional interactions in the brains of adult female Rhesus monkeys with injured and regenerated thoracic spinal cord.
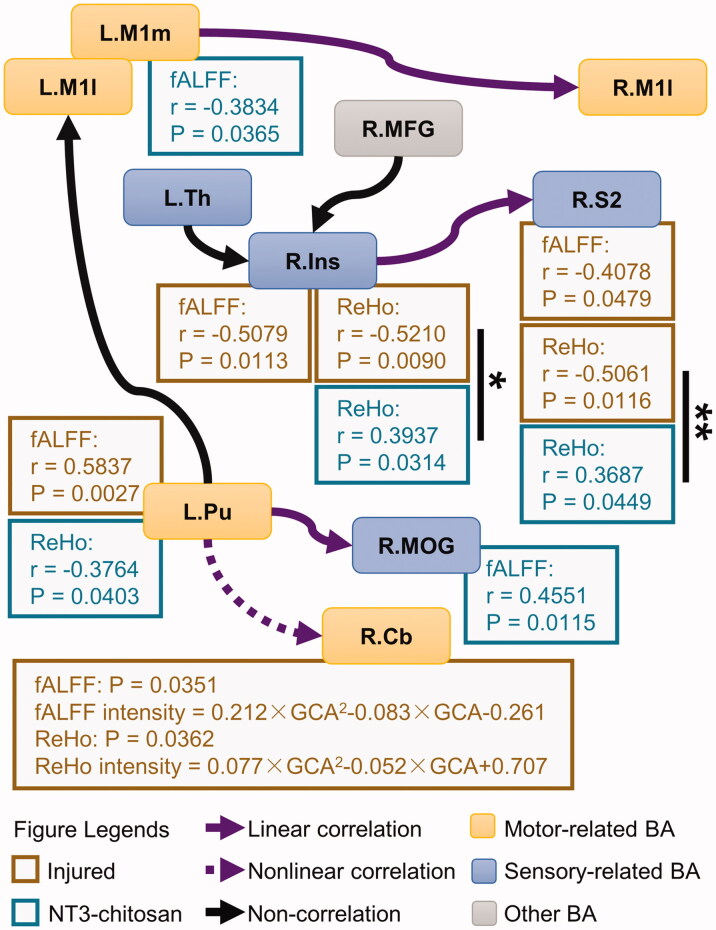
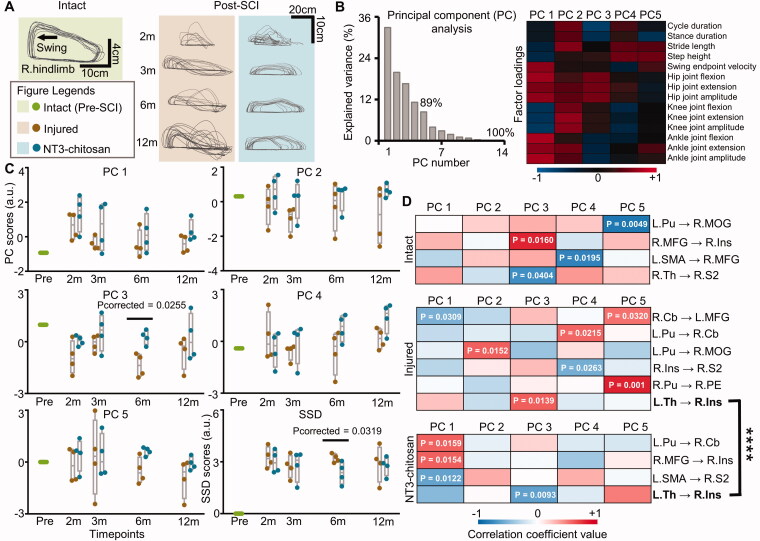
Materials and methods: Resting-state functional magnetic resonance imaging (fMRI) combined with Granger Causality analysis (GCA) and motor behaviour analysis were used to assess the causal interaction between sensorimotor cortices, and calculate the relationship between causal interaction and hindlimb stepping in nine Rhesus monkeys undergoing lesion-induced spontaneous recovery (injured, n = 4) and neurotrophin-3/chitosan transplantation-induced regeneration (NT3-chitosan, n = 5) after SCI.
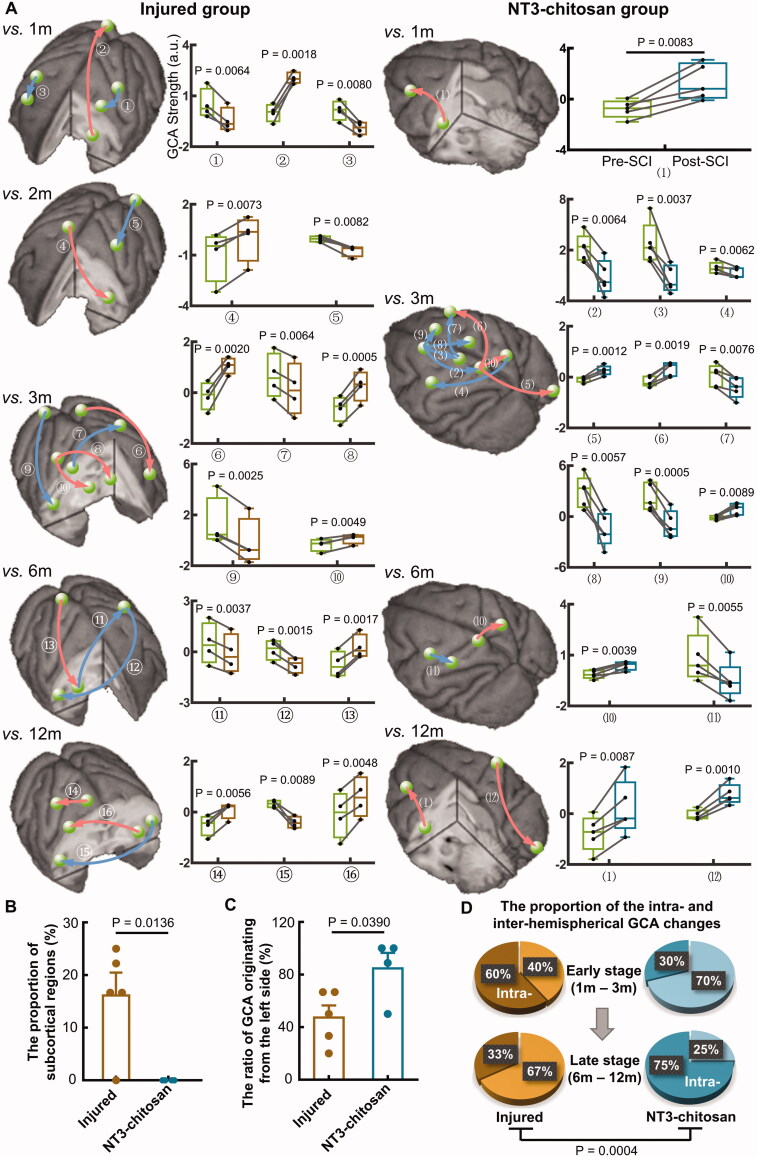
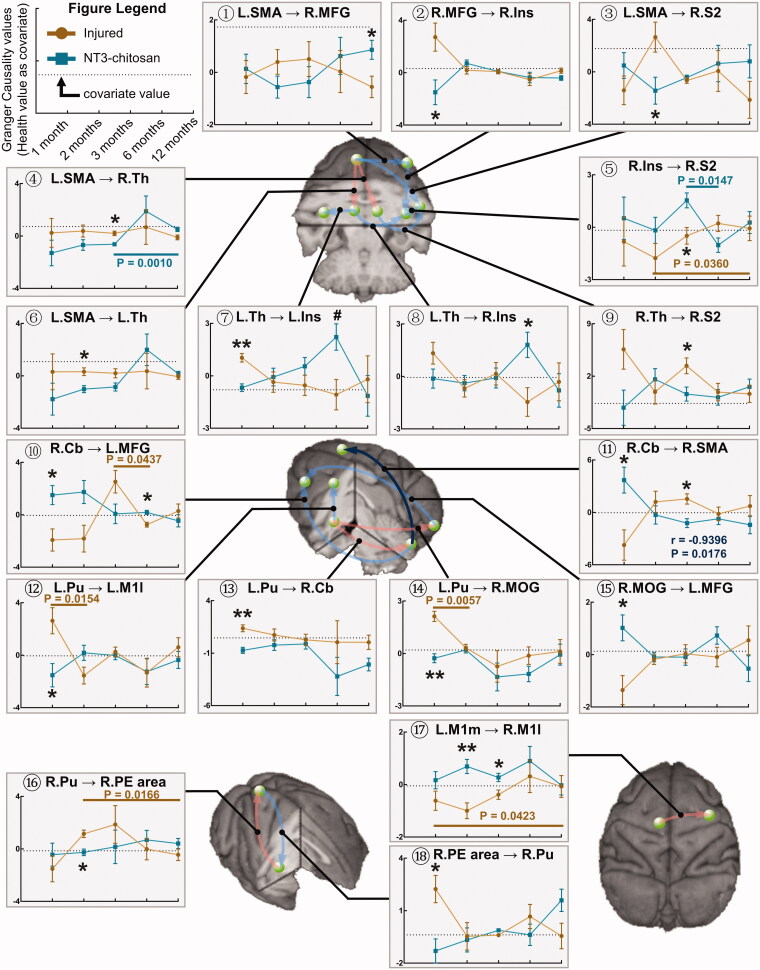
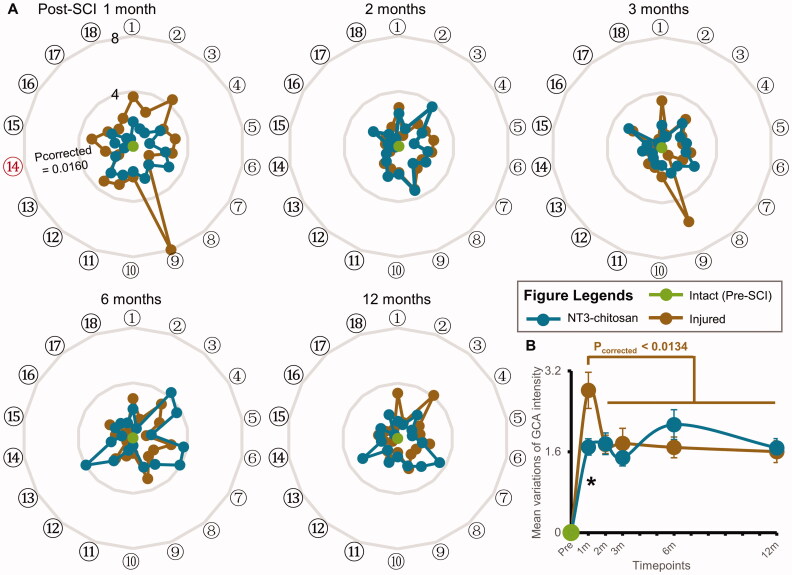
Results: The results showed that the injured and NT3-chitosan-treated animals had distinct spatiotemporal features of brain functional reorganization. The spontaneous recovery followed the model of "early intra-hemispheric reorganization dominant, late inter-hemispheric reorganization dominant", whereas regenerative therapy animals showed the opposite trend. Although the variation degree of information flow intensity was consistent, the tendency and the relationship between local neuronal activity properties and coupling strength were different between the two groups. In addition, the injured and NT3-chitosan-treated animals had similar motor adjustments but various relationship modes between motor performance and information flow intensity.
Conclusions: Our findings show that brain functional reorganization induced by regeneration therapy differed from spontaneous recovery after SCI. The influence of unique changes in brain plasticity on the therapeutic effects of future regeneration therapy strategies should be considered. Key messagesNeural regeneration elicited a unique spatiotemporal mode of brain functional reorganization in the spinal cord injured monkeys, and that regeneration does not simply reverse the process of brain plasticity induced by spinal cord injury (SCI).Independent "properties of local activity - intensity of information flow" relationships between the injured and treated animals indicating that spontaneous recovery and regenerative therapy exerted different effects on the reorganization of the motor network after SCI.A specific information flow from the left thalamus to the right insular can serve as an indicator to reflect a heterogeneous "information flow - motor performance" relationship between injured and treated animals at similar motor adjustments.
Keywords: Nonhuman primate; causal interactions; neural regeneration; reorganization; sensorimotor cortex.
Conflict of interest statement
No potential conflict of interest was reported by the authors.
Figures
References
-
- Ghosh A, Haiss F, Sydekum E, et al. Rewiring of hindlimb corticospinal neurons after spinal cord injury. Nat Neurosci. 2010;13(1):97–104. - PubMed
-
- Sydekum E, Ghosh A, Gullo M, et al. Rapid functional reorganization of the forelimb cortical representation after thoracic spinal cord injury in adult rats. NeuroImage. 2014;87:72–79. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical