

A single-cell analysis of thymopoiesis and thymic iNKT cell development in pigs
- PMID: 35793622
- PMCID: PMC9704770
- DOI: 10.1016/j.celrep.2022.111050
A single-cell analysis of thymopoiesis and thymic iNKT cell development in pigs
Abstract
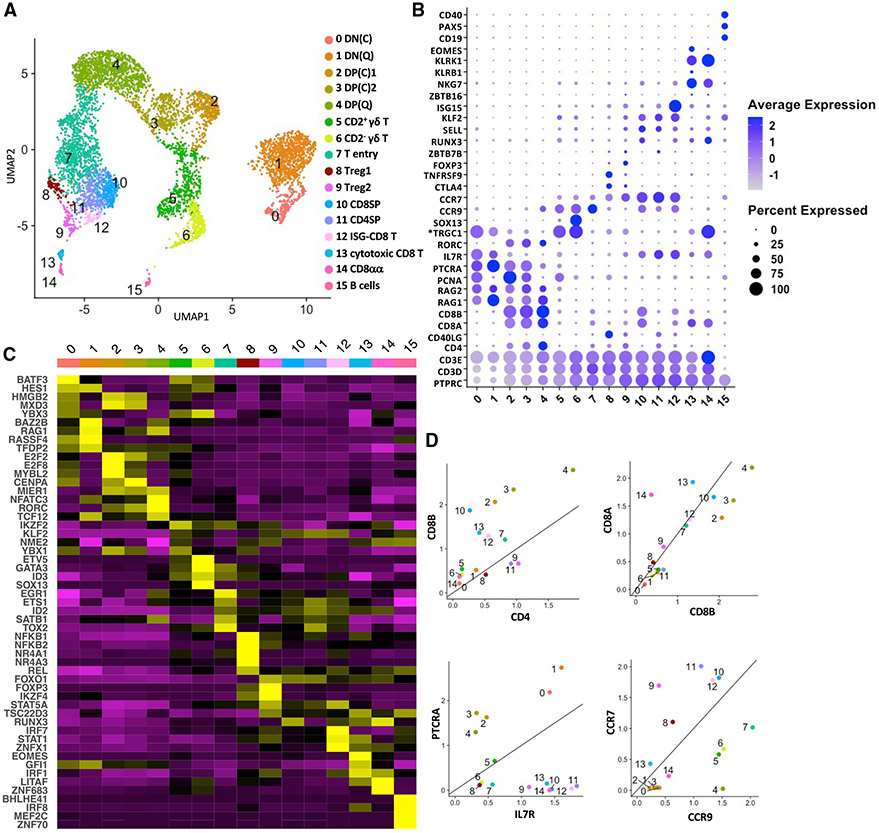
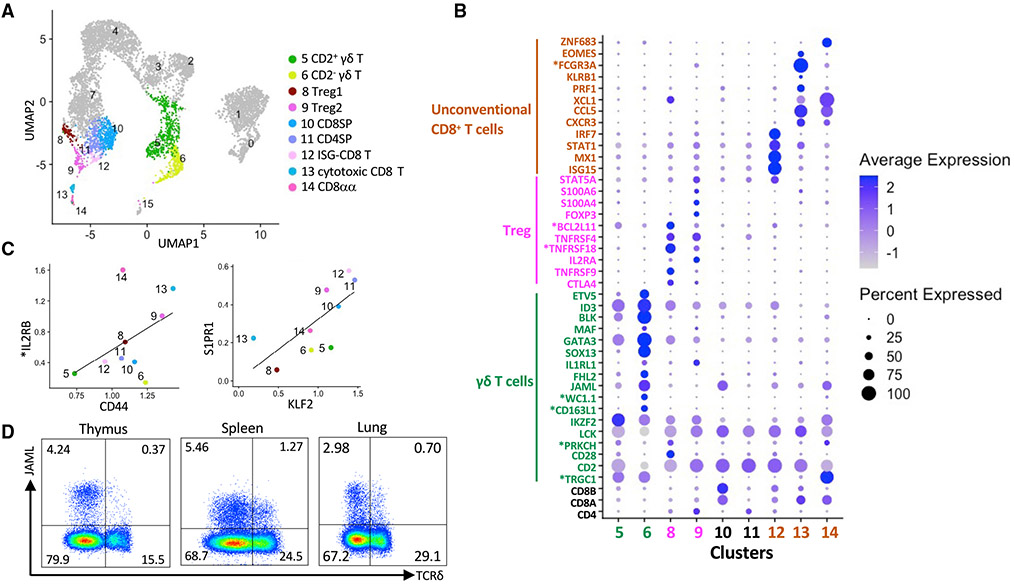
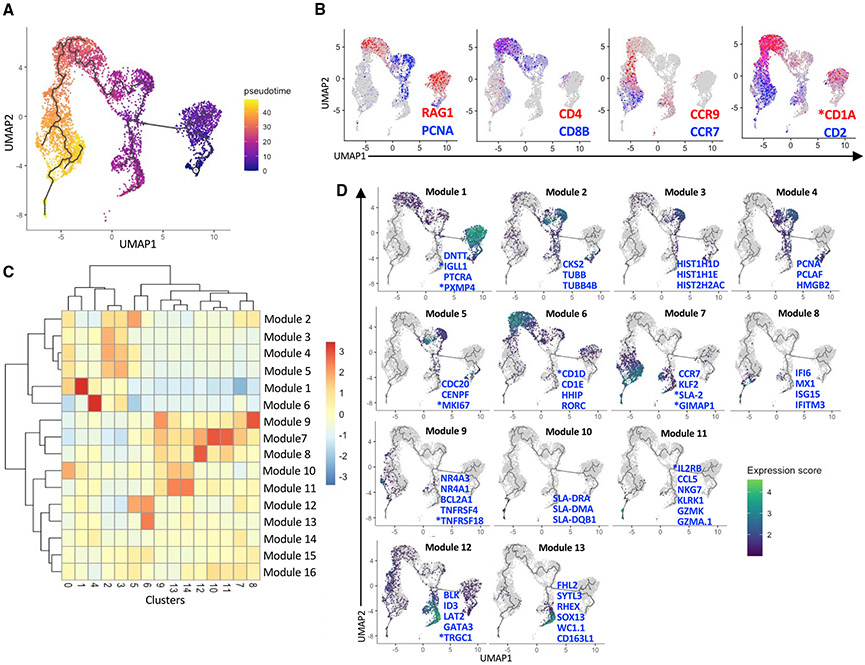
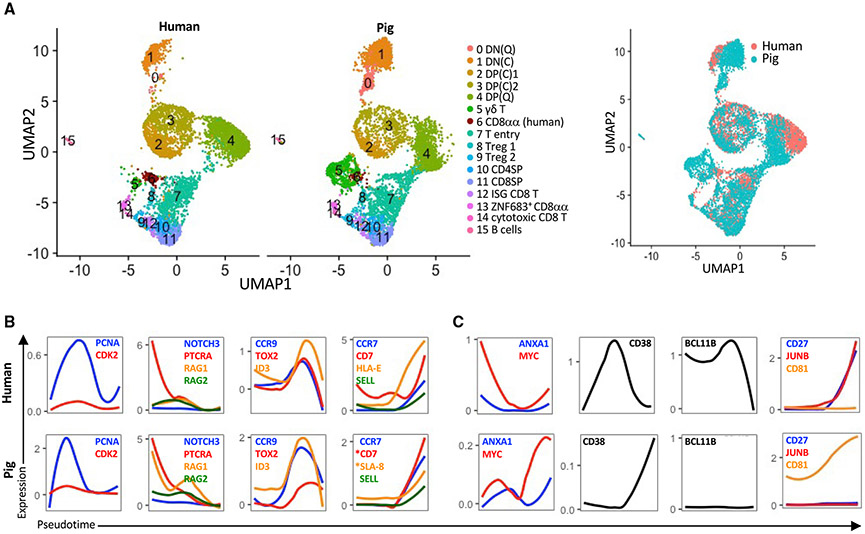
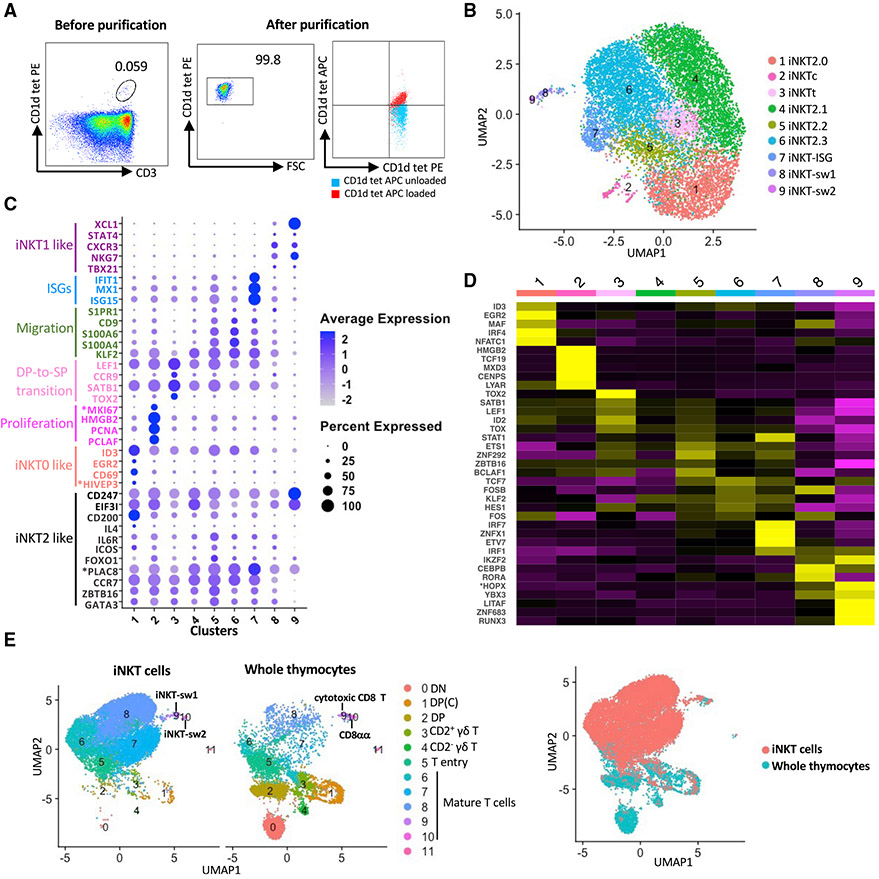
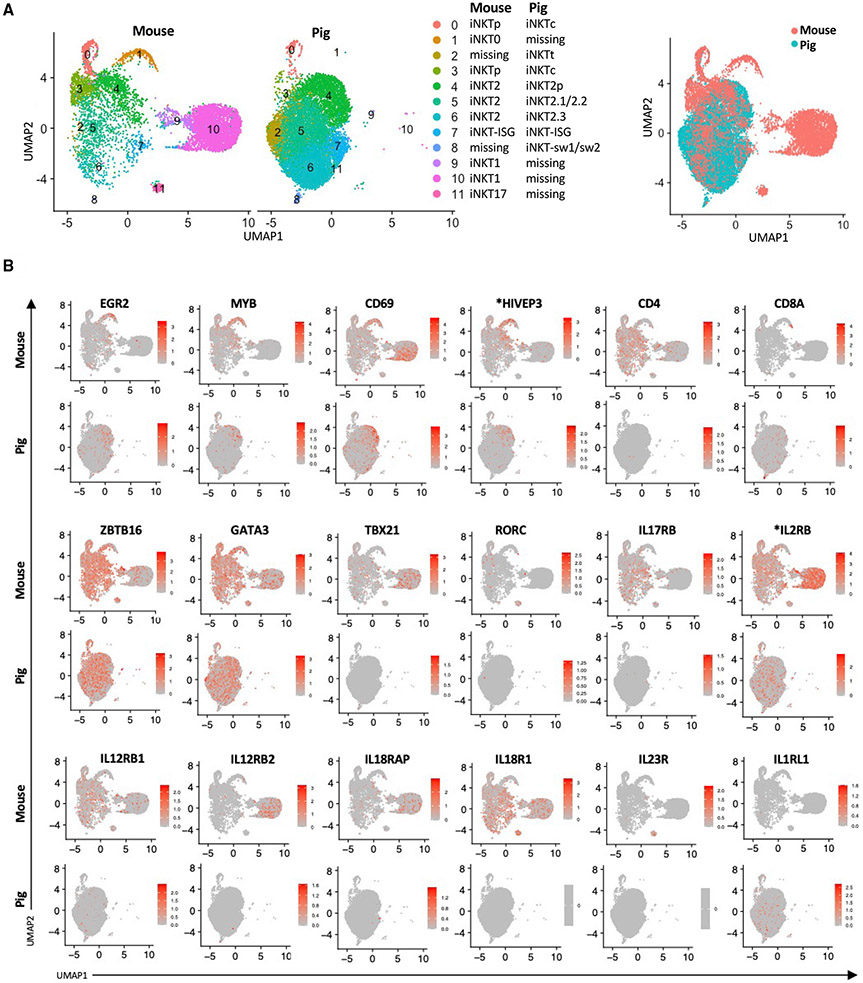
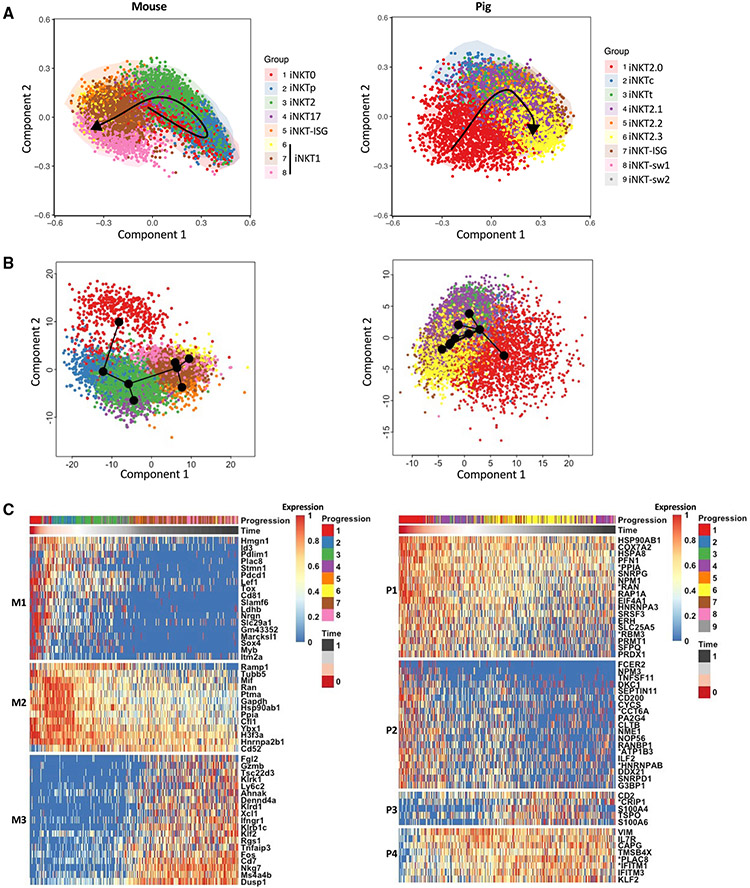
Many aspects of the porcine immune system remain poorly characterized, which poses a barrier to improving swine health and utilizing pigs as preclinical models. Here, we employ single-cell RNA sequencing (scRNA-seq) to create a cell atlas of the early-adolescent pig thymus. Our data show conserved features as well as species-specific differences in cell states and cell types compared with human thymocytes. We also describe several unconventional T cell types with gene expression profiles associated with innate effector functions. This includes a cell census of more than 11,000 differentiating invariant natural killer T (iNKT) cells, which reveals that the functional diversity of pig iNKT cells differs substantially from the iNKT0/1/2/17 subset differentiation paradigm established in mice. Our data characterize key differentiation events in porcine thymopoiesis and iNKT cell maturation and provide important insights into pig T cell development.
Keywords: CP: Immunology; invariant natural killer T cell; porcine; single cell transcriptomics; thymopoiesis.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Baranek T, Lebrigand K, de Amat Herbozo C, Gonzalez L, Bogard G, Dietrich C, Magnone V, Boisseau C, Jouan Y, Trottein F, et al. (2020). High dimensional single-cell analysis reveals iNKT cell developmental trajectories and effector fate decision. Cell Rep. 32, 108116. 10.1016/j.celrep.2020.108116. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
