

Neural network organization for courtship-song feature detection in Drosophila
- PMID: 35793679
- PMCID: PMC9378594
- DOI: 10.1016/j.cub.2022.06.019
Neural network organization for courtship-song feature detection in Drosophila
Abstract
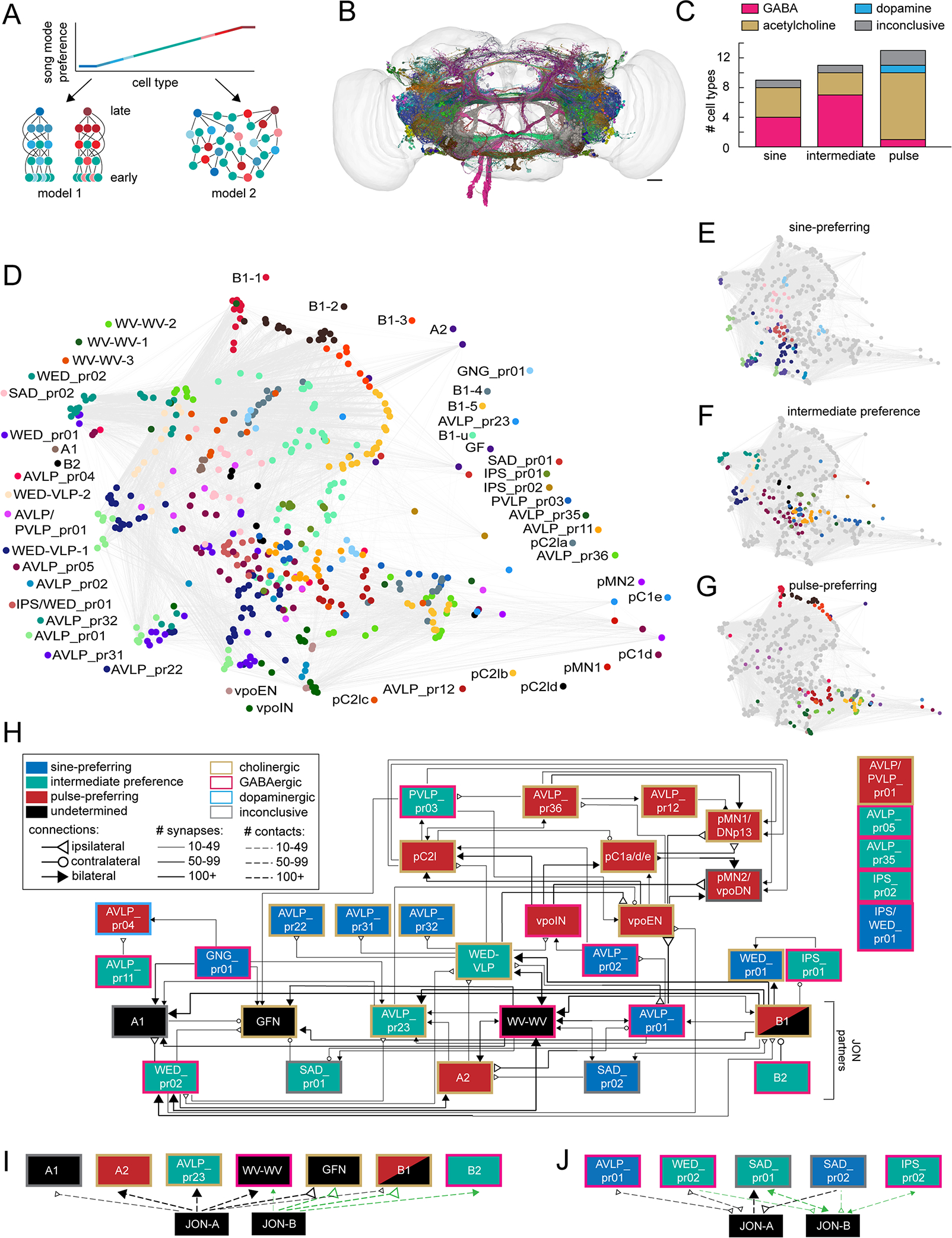
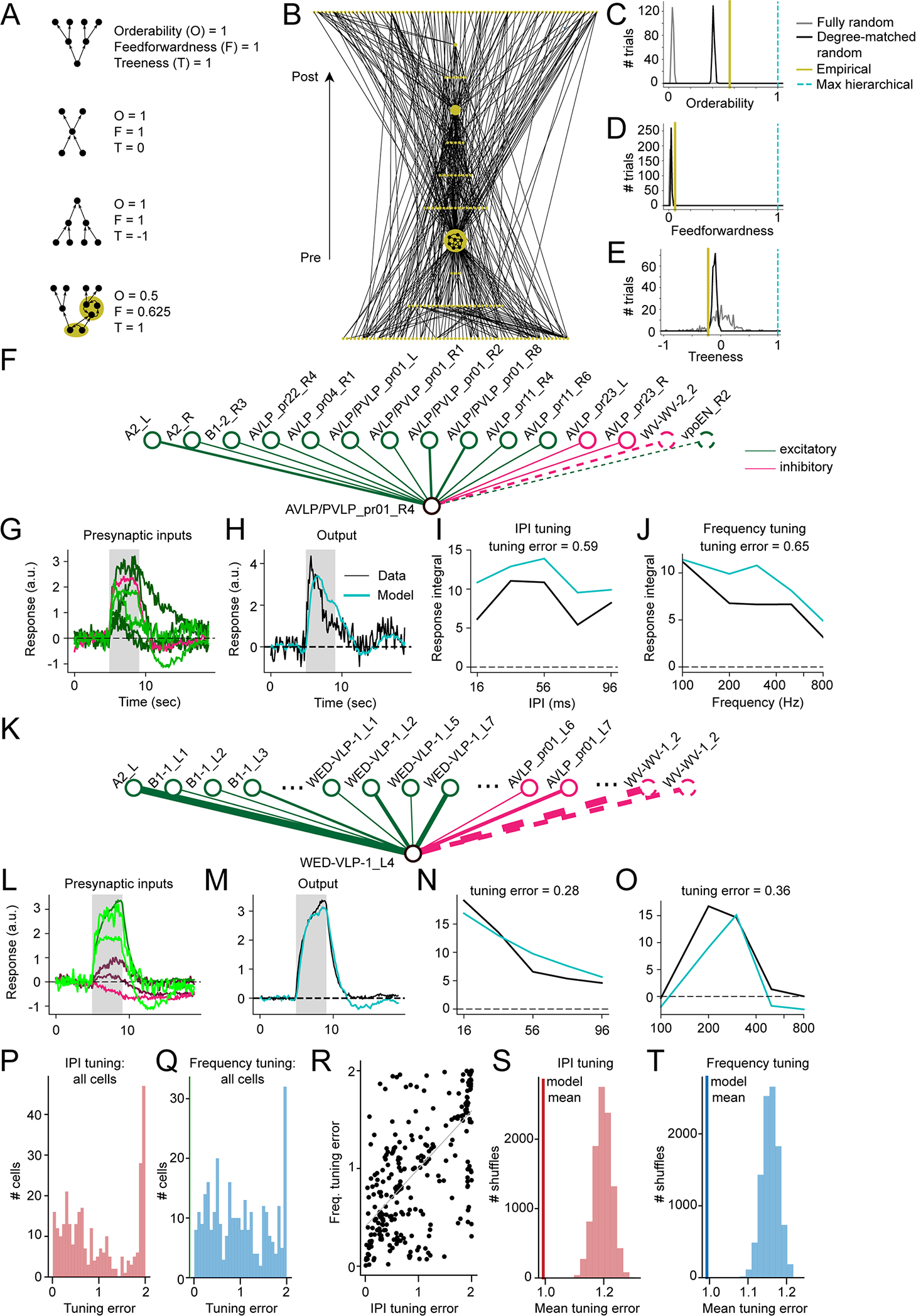
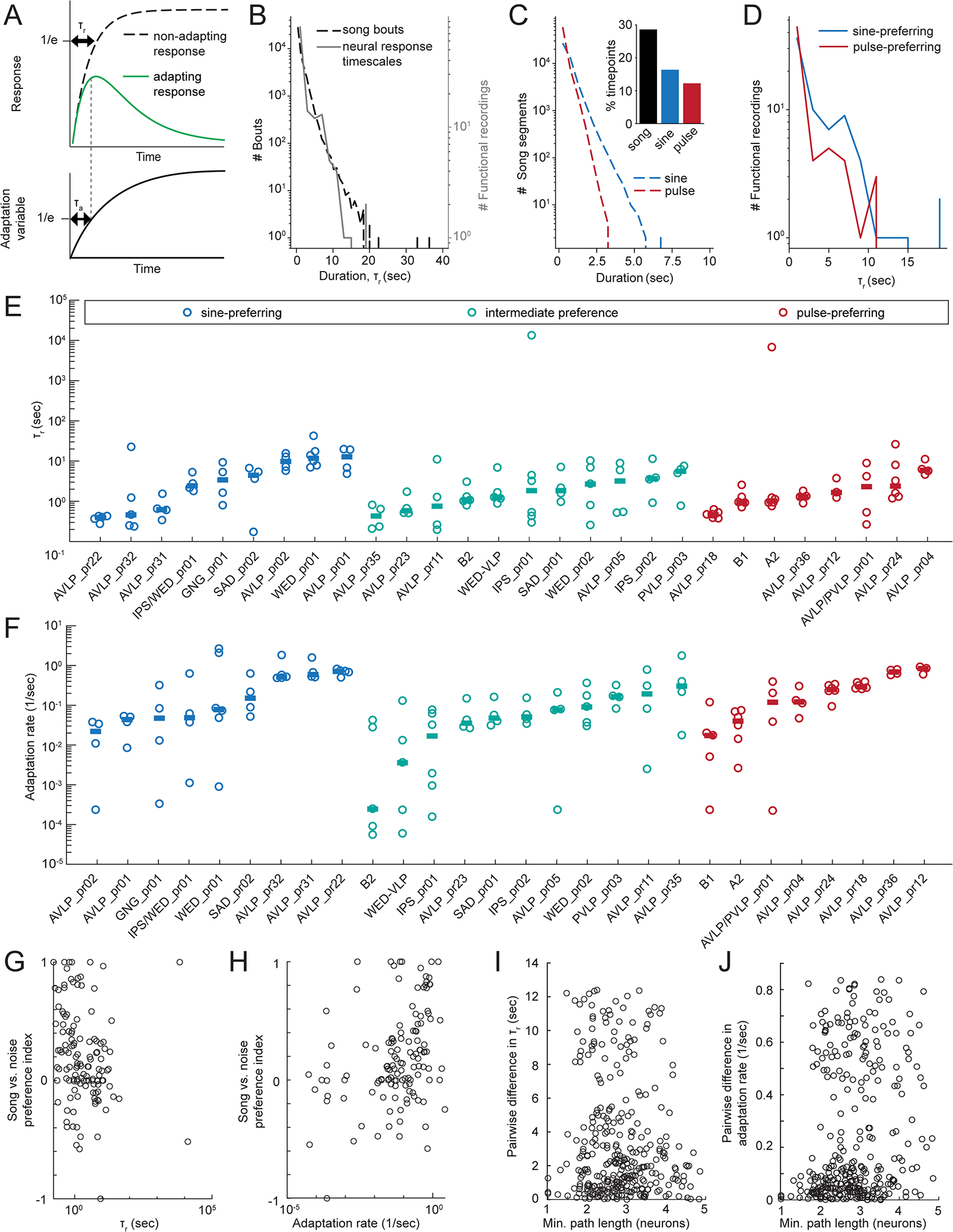
Animals communicate using sounds in a wide range of contexts, and auditory systems must encode behaviorally relevant acoustic features to drive appropriate reactions. How feature detection emerges along auditory pathways has been difficult to solve due to challenges in mapping the underlying circuits and characterizing responses to behaviorally relevant features. Here, we study auditory activity in the Drosophila melanogaster brain and investigate feature selectivity for the two main modes of fly courtship song, sinusoids and pulse trains. We identify 24 new cell types of the intermediate layers of the auditory pathway, and using a new connectomic resource, FlyWire, we map all synaptic connections between these cell types, in addition to connections to known early and higher-order auditory neurons-this represents the first circuit-level map of the auditory pathway. We additionally determine the sign (excitatory or inhibitory) of most synapses in this auditory connectome. We find that auditory neurons display a continuum of preferences for courtship song modes and that neurons with different song-mode preferences and response timescales are highly interconnected in a network that lacks hierarchical structure. Nonetheless, we find that the response properties of individual cell types within the connectome are predictable from their inputs. Our study thus provides new insights into the organization of auditory coding within the Drosophila brain.
Keywords: acoustic communication; auditory; calcium imaging; connectomics; neural network; sensory responses.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
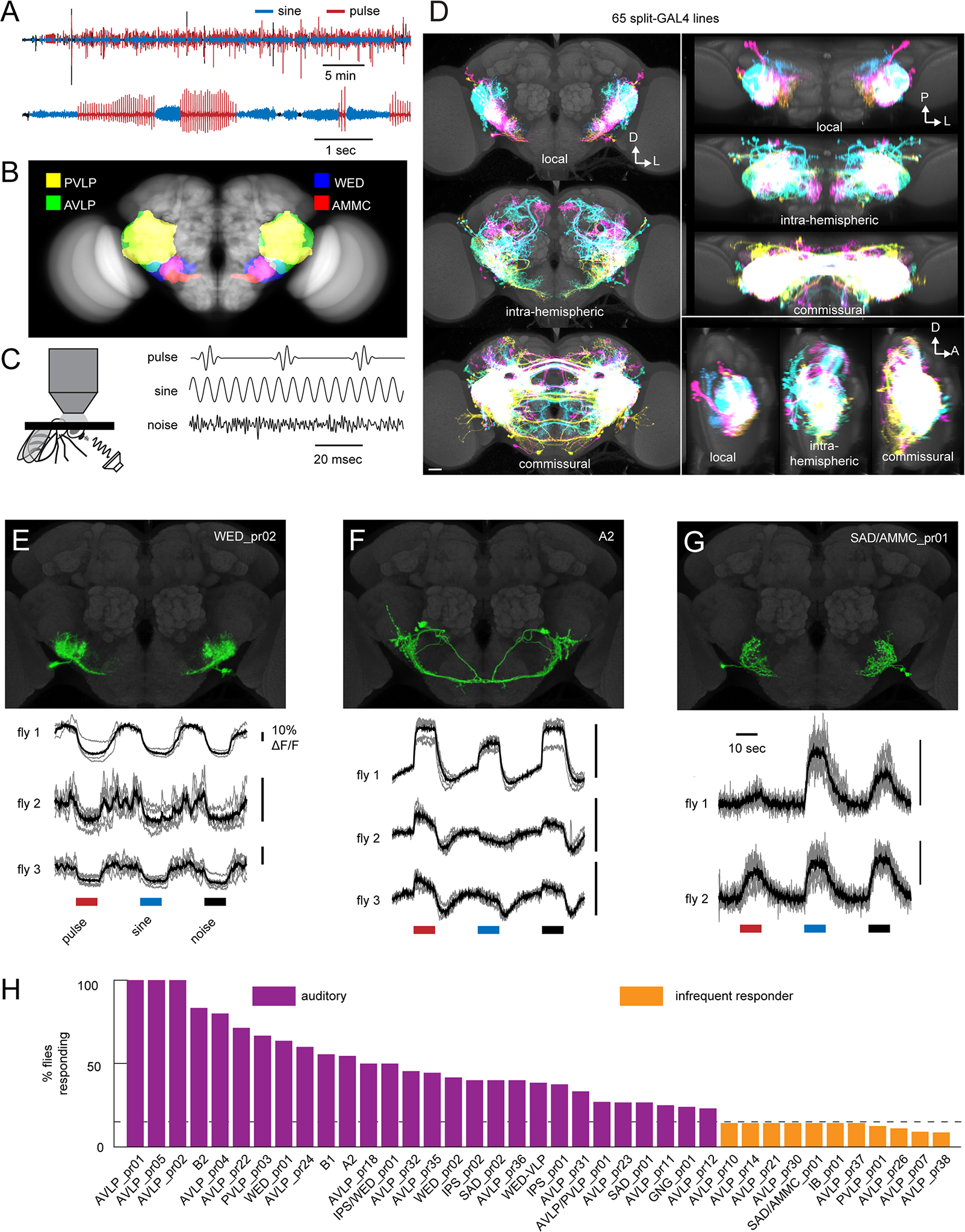
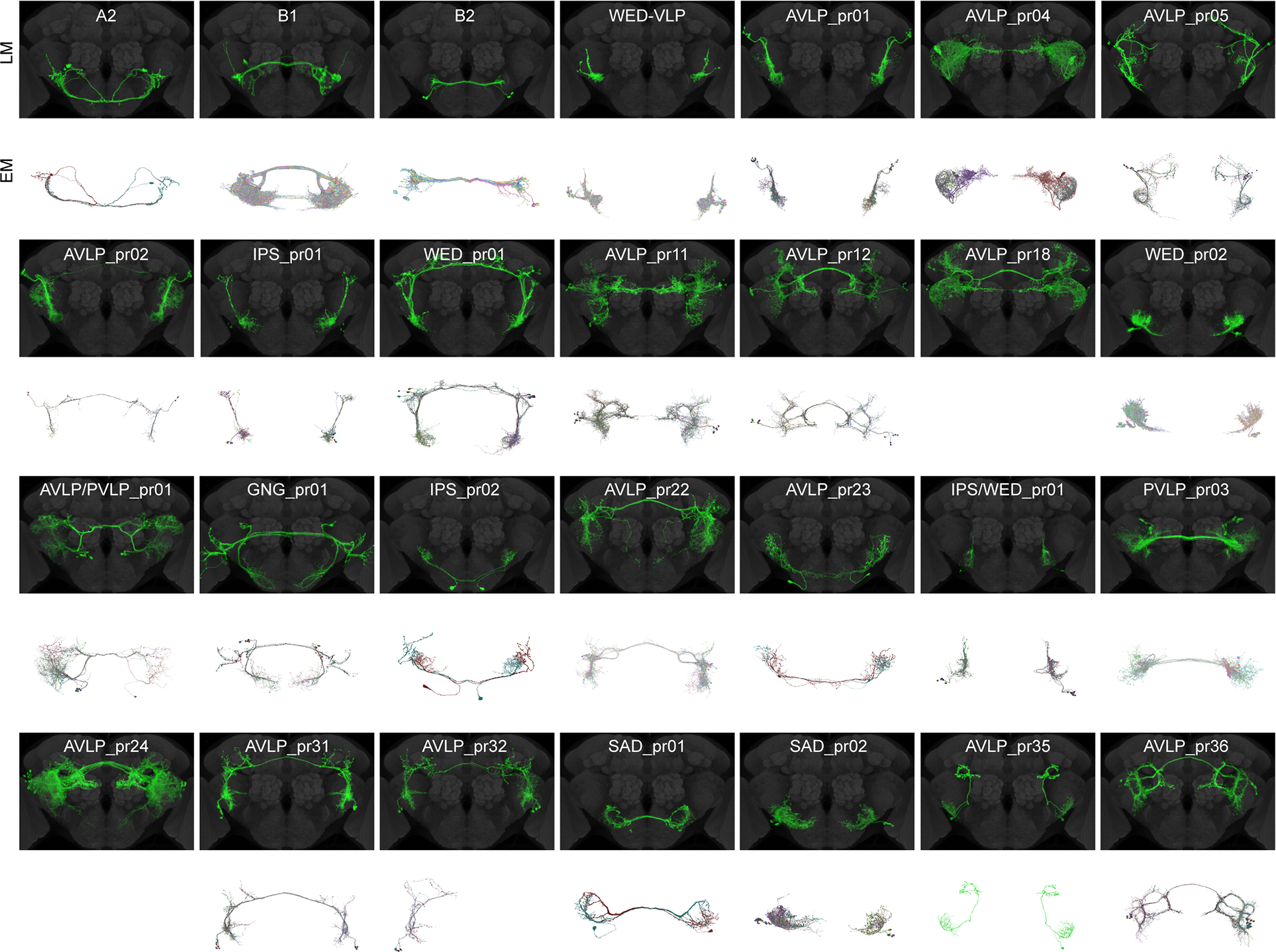
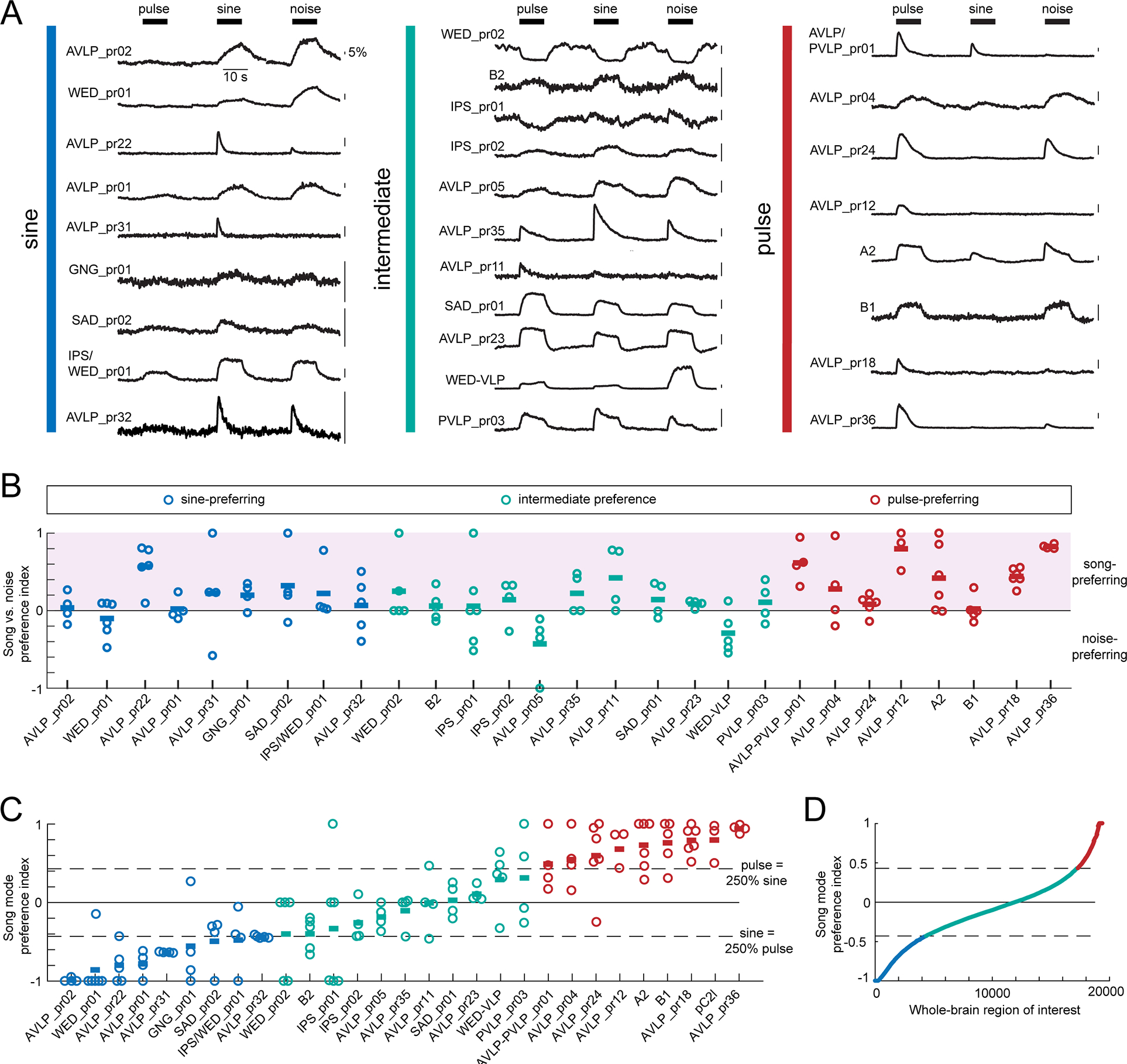
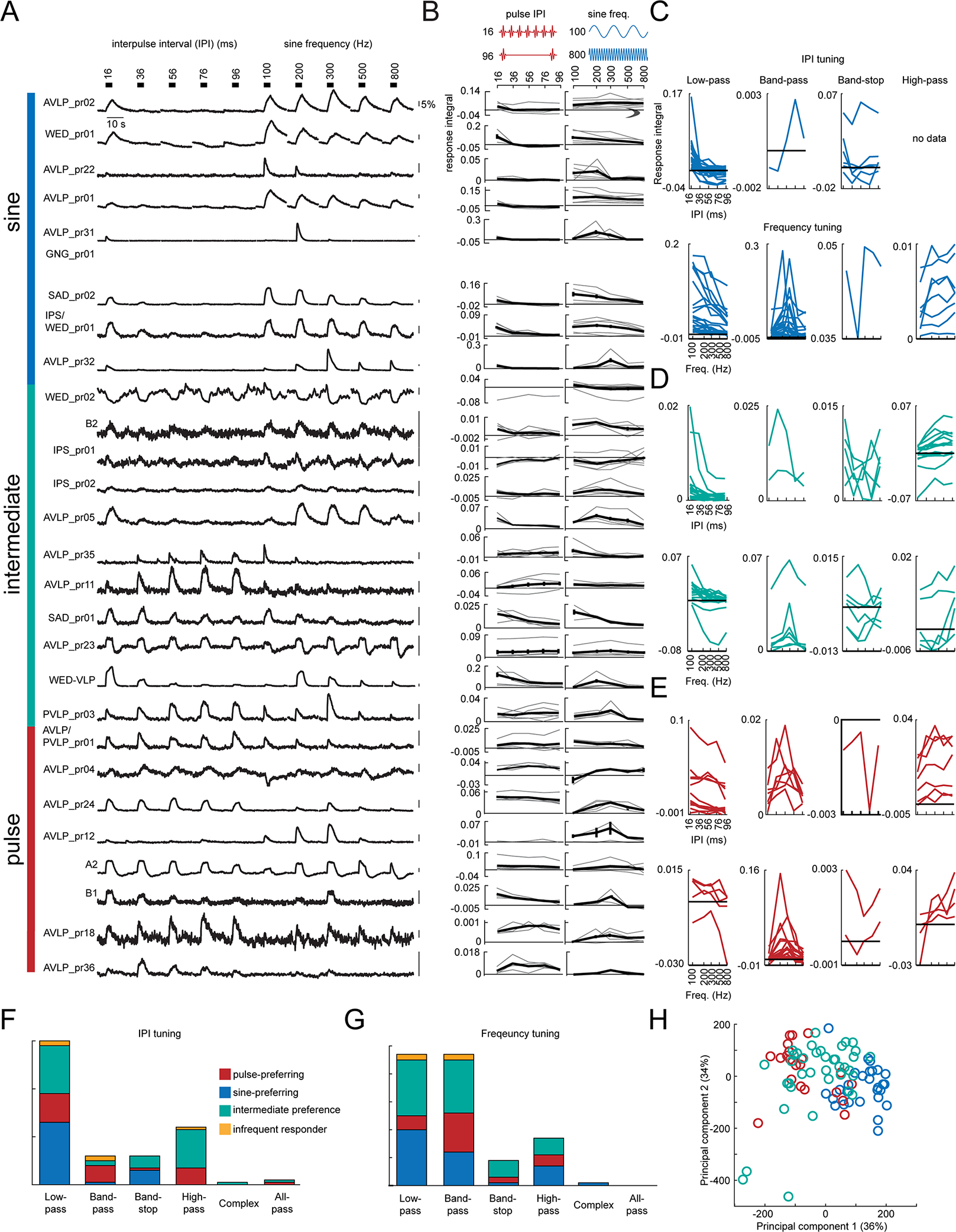
Comment in
-
Systems neuroscience: Auditory processing at synaptic resolution.Curr Biol. 2022 Aug 8;32(15):R830-R833. doi: 10.1016/j.cub.2022.07.001. Curr Biol. 2022. PMID: 35944481
References
-
- Behr O, and von Helversen O (2004). Bat serenades—complex courtship songs of the sac-winged bat (Saccopteryx bilineata). Behavioral Ecology and Sociobiology 56, 106–115.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
