

Atypical cyclic di-AMP signaling is essential for Porphyromonas gingivalis growth and regulation of cell envelope homeostasis and virulence
- PMID: 35794154
- PMCID: PMC9259658
- DOI: 10.1038/s41522-022-00316-w
Atypical cyclic di-AMP signaling is essential for Porphyromonas gingivalis growth and regulation of cell envelope homeostasis and virulence
Abstract
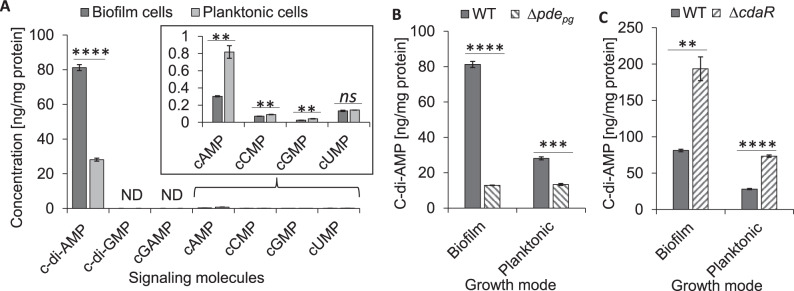
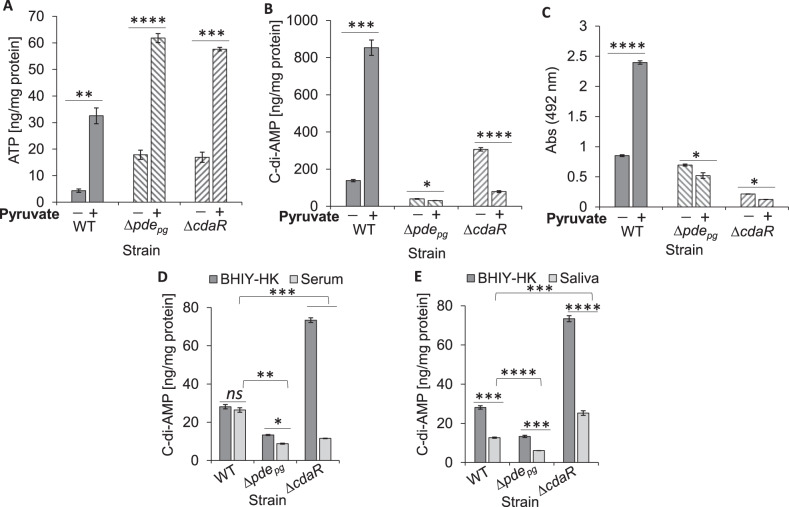
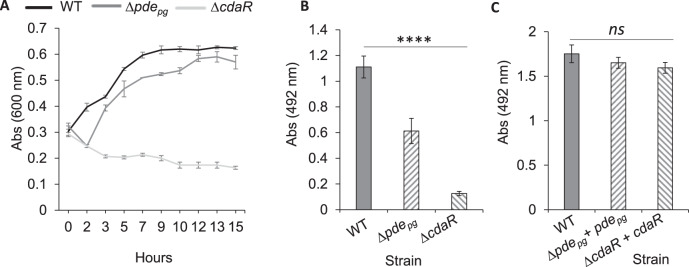
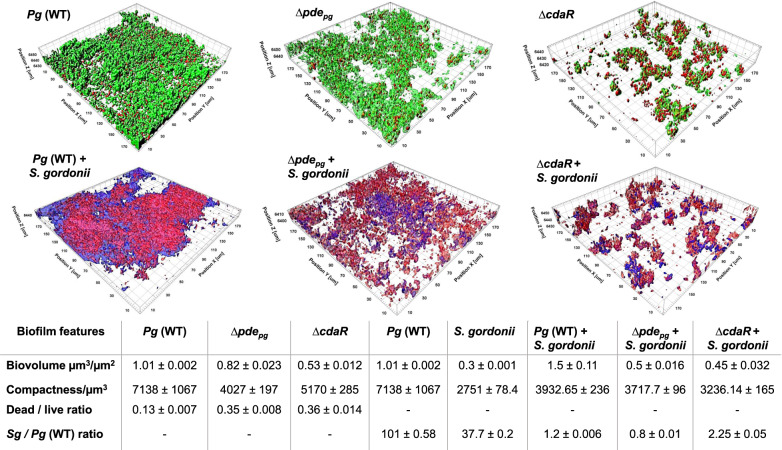
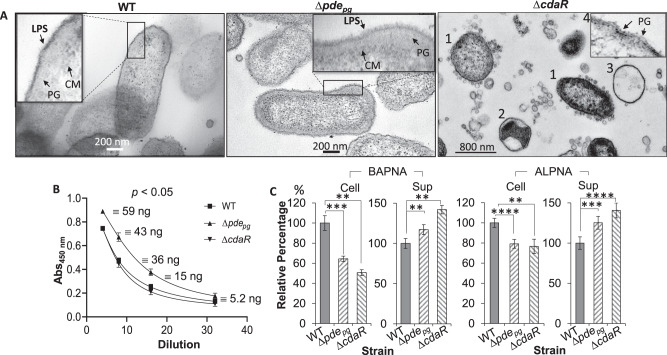
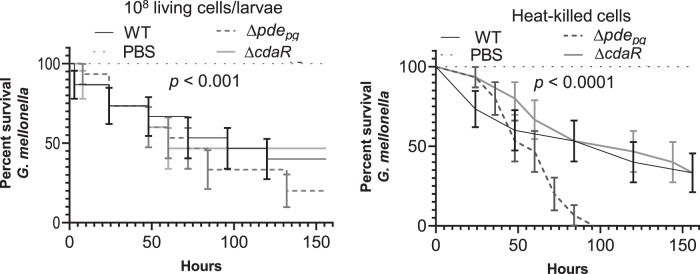
Microbial pathogens employ signaling systems through cyclic (di-) nucleotide monophosphates serving as second messengers to increase fitness during pathogenesis. However, signaling schemes via second messengers in Porphyromonas gingivalis, a key Gram-negative anaerobic oral pathogen, remain unknown. Here, we report that among various ubiquitous second messengers, P. gingivalis strains predominantly synthesize bis-(3',5')-cyclic di-adenosine monophosphate (c-di-AMP), which is essential for their growth and survival. Our findings demonstrate an unusual regulation of c-di-AMP synthesis in P. gingivalis. P. gingivalis c-di-AMP phosphodiesterase (PDE) gene (pdepg) positively regulates c-di-AMP synthesis and impedes a decrease in c-di-AMP concentration despite encoding conserved amino acid motifs for phosphodiesterase activity. Instead, the predicted regulator gene cdaR, unrelated to the c-di-AMP PDE genes, serves as a potent negative regulator of c-di-AMP synthesis in this anaerobe. Further, our findings reveal that pdepg and cdaR are required to regulate the incorporation of ATP into c-di-AMP upon pyruvate utilization, leading to enhanced biofilm formation. We show that shifts in c-di-AMP signaling change the integrity and homeostasis of cell envelope, importantly, the structure and immunoreactivity of the lipopolysaccharide layer. Additionally, microbe-microbe interactions and the virulence potential of P. gingivalis were modulated by c-di-AMP. These studies provide the first glimpse into the scheme of second messenger signaling in P. gingivalis and perhaps other Bacteroidetes. Further, our findings indicate that c-di-AMP signaling promotes the fitness of the residents of the oral cavity and the development of a pathogenic community.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
The multifaceted role of c-di-AMP signaling in the regulation of Porphyromonas gingivalis lipopolysaccharide structure and function.Front Cell Infect Microbiol. 2024 Jun 12;14:1418651. doi: 10.3389/fcimb.2024.1418651. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38933693 Free PMC article.
-
Cyclic di-AMP mediates biofilm formation.Mol Microbiol. 2016 Mar;99(5):945-59. doi: 10.1111/mmi.13277. Epub 2015 Dec 15. Mol Microbiol. 2016. PMID: 26564551 Free PMC article.
-
An Essential Poison: Synthesis and Degradation of Cyclic Di-AMP in Bacillus subtilis.J Bacteriol. 2015 Oct;197(20):3265-74. doi: 10.1128/JB.00564-15. Epub 2015 Aug 3. J Bacteriol. 2015. PMID: 26240071 Free PMC article.
-
A jack of all trades: the multiple roles of the unique essential second messenger cyclic di-AMP.Mol Microbiol. 2015 Jul;97(2):189-204. doi: 10.1111/mmi.13026. Epub 2015 May 9. Mol Microbiol. 2015. PMID: 25869574 Review.
-
Replenishing the cyclic-di-AMP pool: regulation of diadenylate cyclase activity in bacteria.Curr Genet. 2016 Nov;62(4):731-738. doi: 10.1007/s00294-016-0600-8. Epub 2016 Apr 13. Curr Genet. 2016. PMID: 27074767 Review.
Cited by
-
Unveiling the molecular mechanisms of the type IX secretion system's response regulator: Structural and functional insights.PNAS Nexus. 2024 Jul 31;3(8):pgae316. doi: 10.1093/pnasnexus/pgae316. eCollection 2024 Aug. PNAS Nexus. 2024. PMID: 39139265 Free PMC article.
-
Is biofilm formation intrinsic to the origin of life?Environ Microbiol. 2023 Jan;25(1):26-39. doi: 10.1111/1462-2920.16179. Epub 2022 Sep 7. Environ Microbiol. 2023. PMID: 36655713 Free PMC article.
-
Near-infrared light-triggered nitric oxide nanocomposites for photodynamic/photothermal complementary therapy against periodontal biofilm in an animal model.Theranostics. 2023 Apr 17;13(7):2350-2367. doi: 10.7150/thno.83745. eCollection 2023. Theranostics. 2023. PMID: 37153739 Free PMC article.
-
Design and application of antimicrobial nanomaterials in the treatment of periodontitis.Nanomedicine (Lond). 2025 Apr;20(7):707-723. doi: 10.1080/17435889.2025.2469492. Epub 2025 Mar 5. Nanomedicine (Lond). 2025. PMID: 40042364 Review.
-
Characterization of c-di-AMP signaling in the periodontal pathobiont, Treponema denticola.Mol Oral Microbiol. 2024 Oct;39(5):354-367. doi: 10.1111/omi.12458. Epub 2024 Mar 4. Mol Oral Microbiol. 2024. PMID: 38436552
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
