

Molecular landscapes of human hippocampal immature neurons across lifespan
- PMID: 35794479
- PMCID: PMC9316413
- DOI: 10.1038/s41586-022-04912-w
Molecular landscapes of human hippocampal immature neurons across lifespan
Abstract
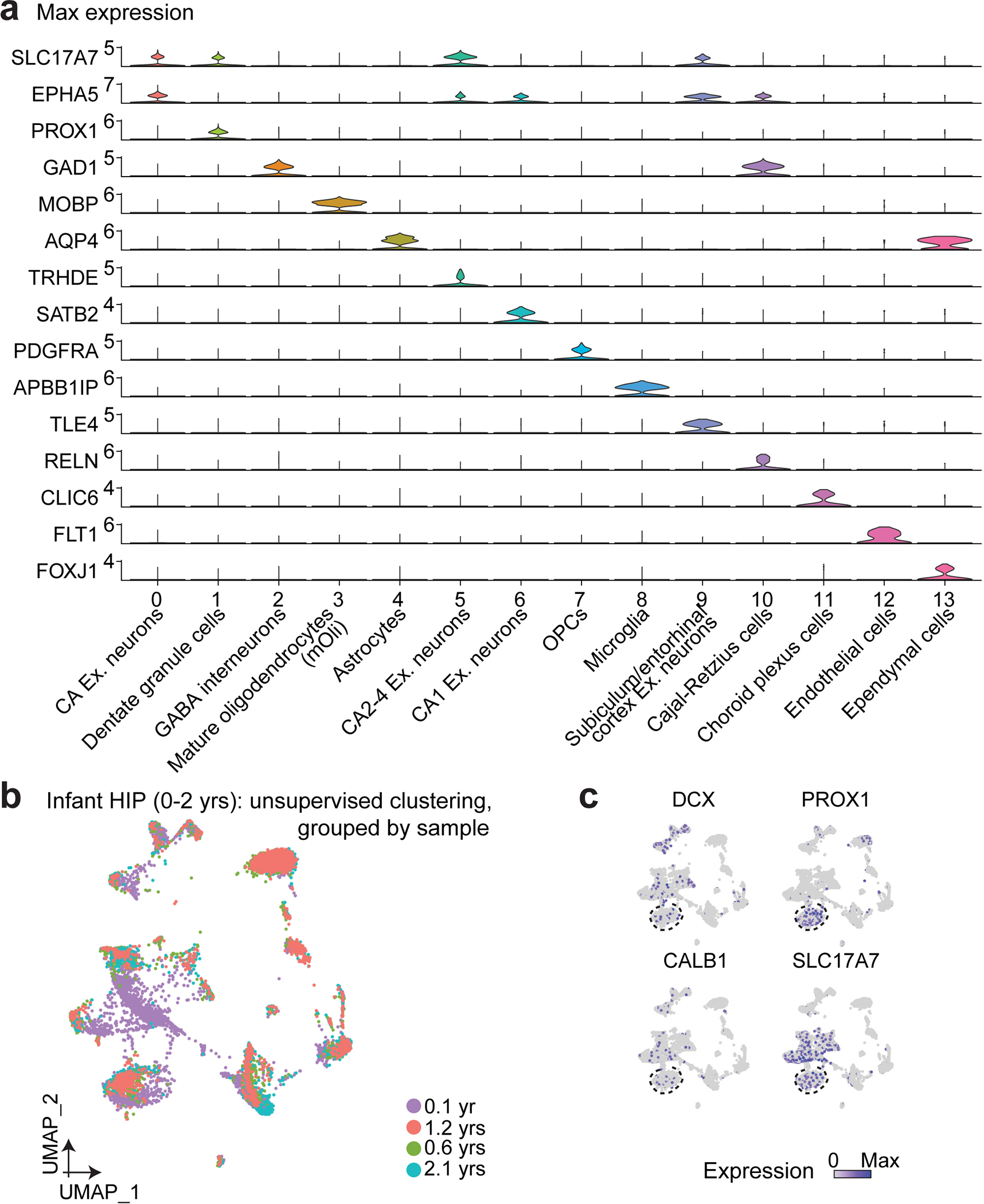
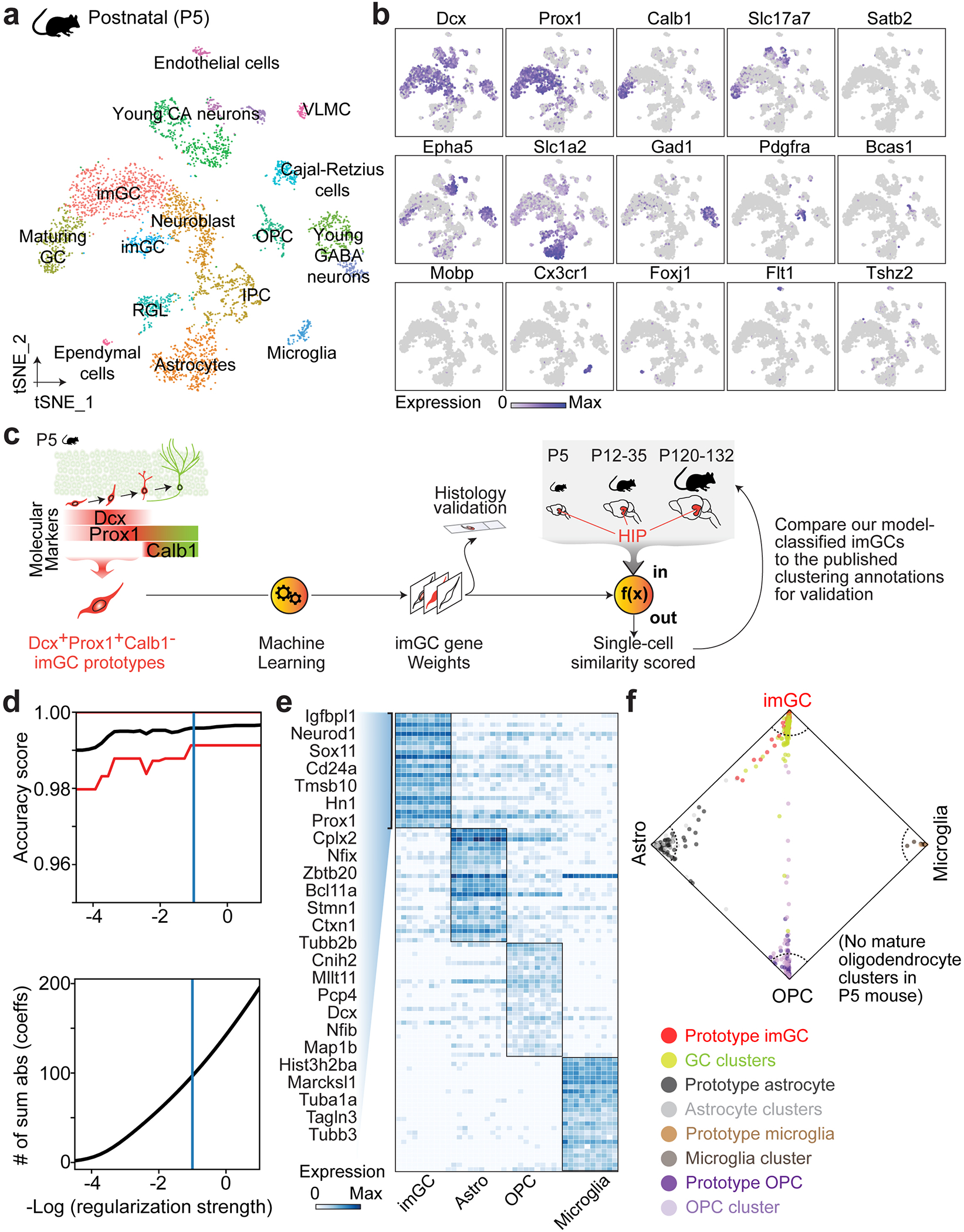
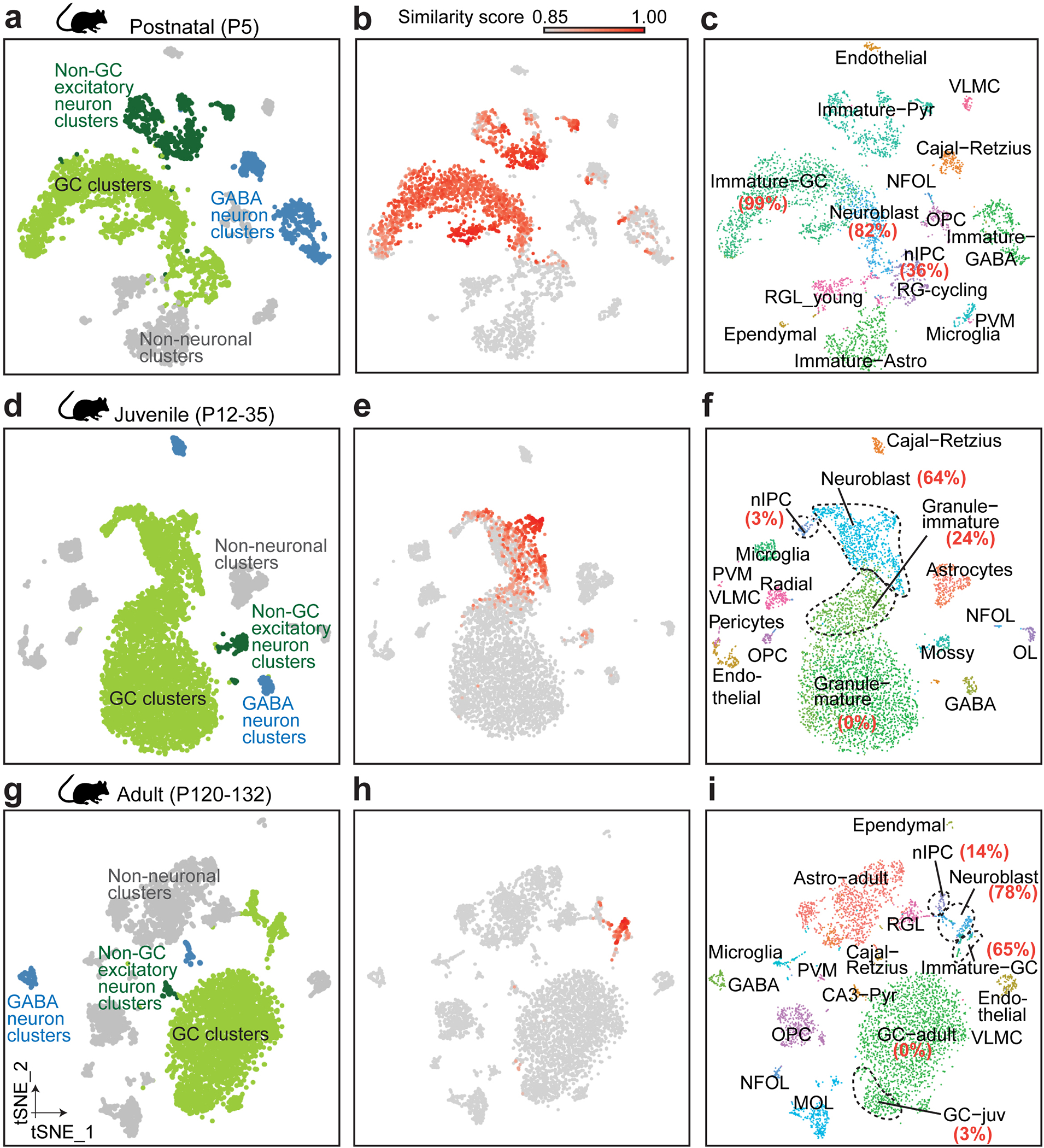
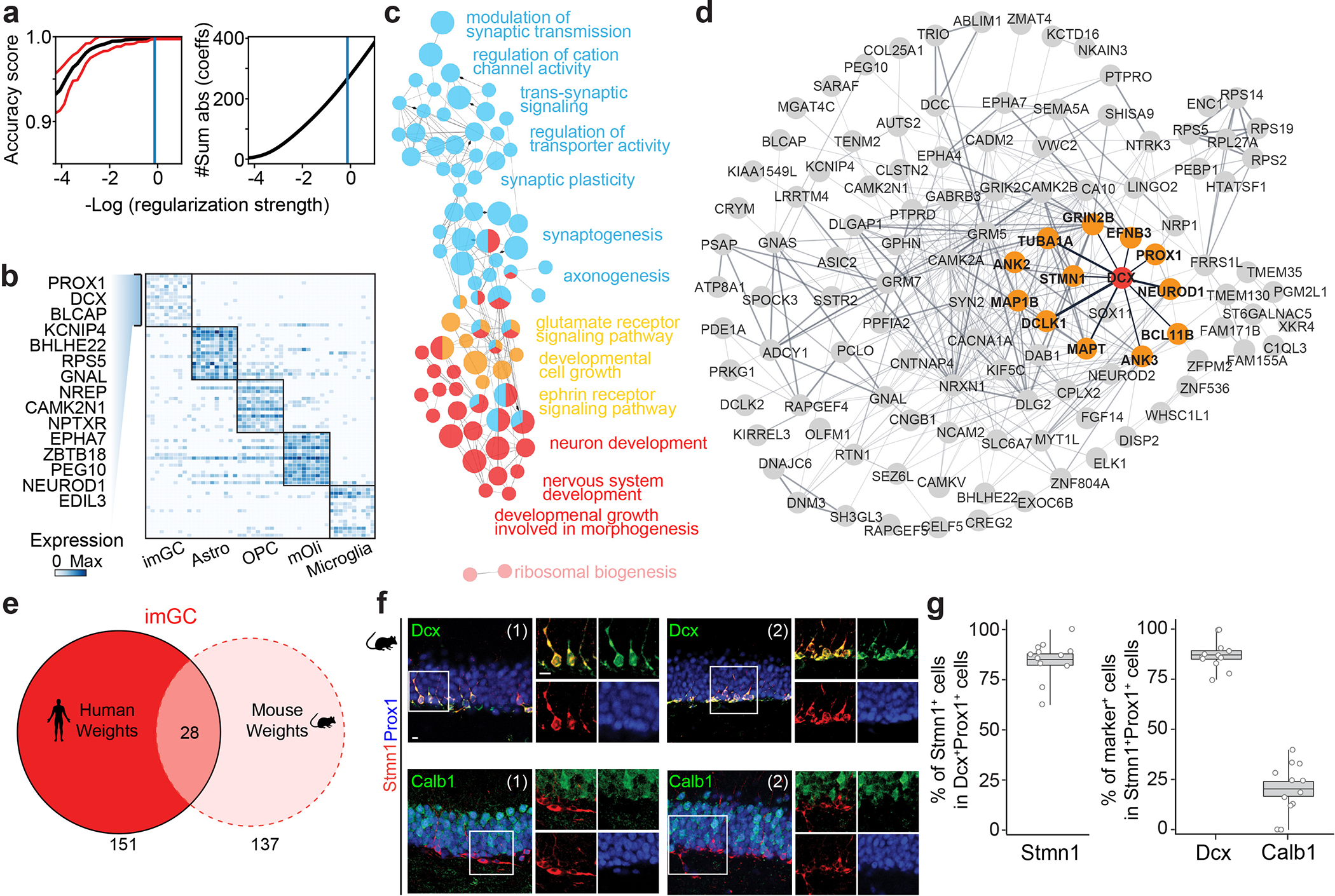
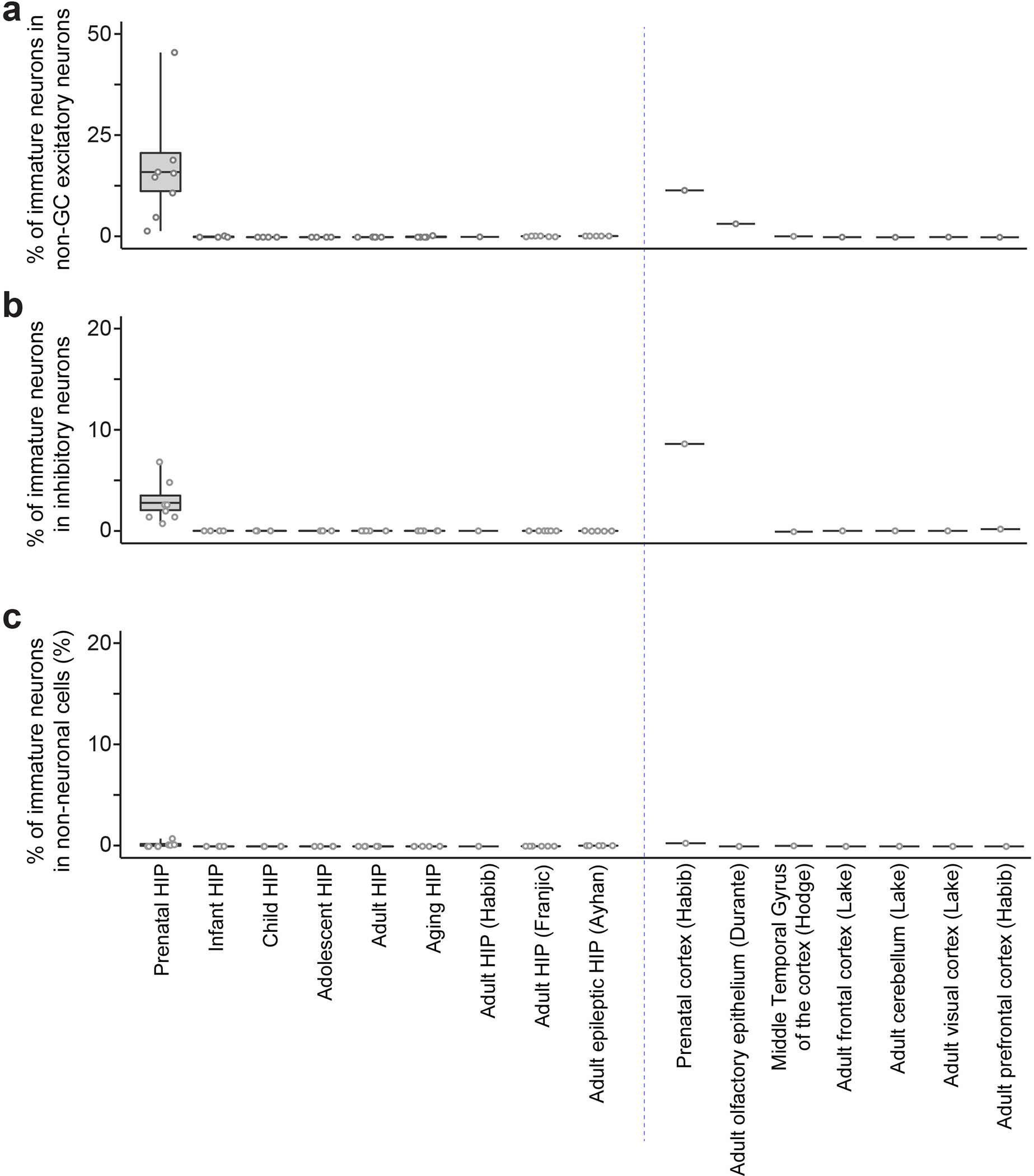
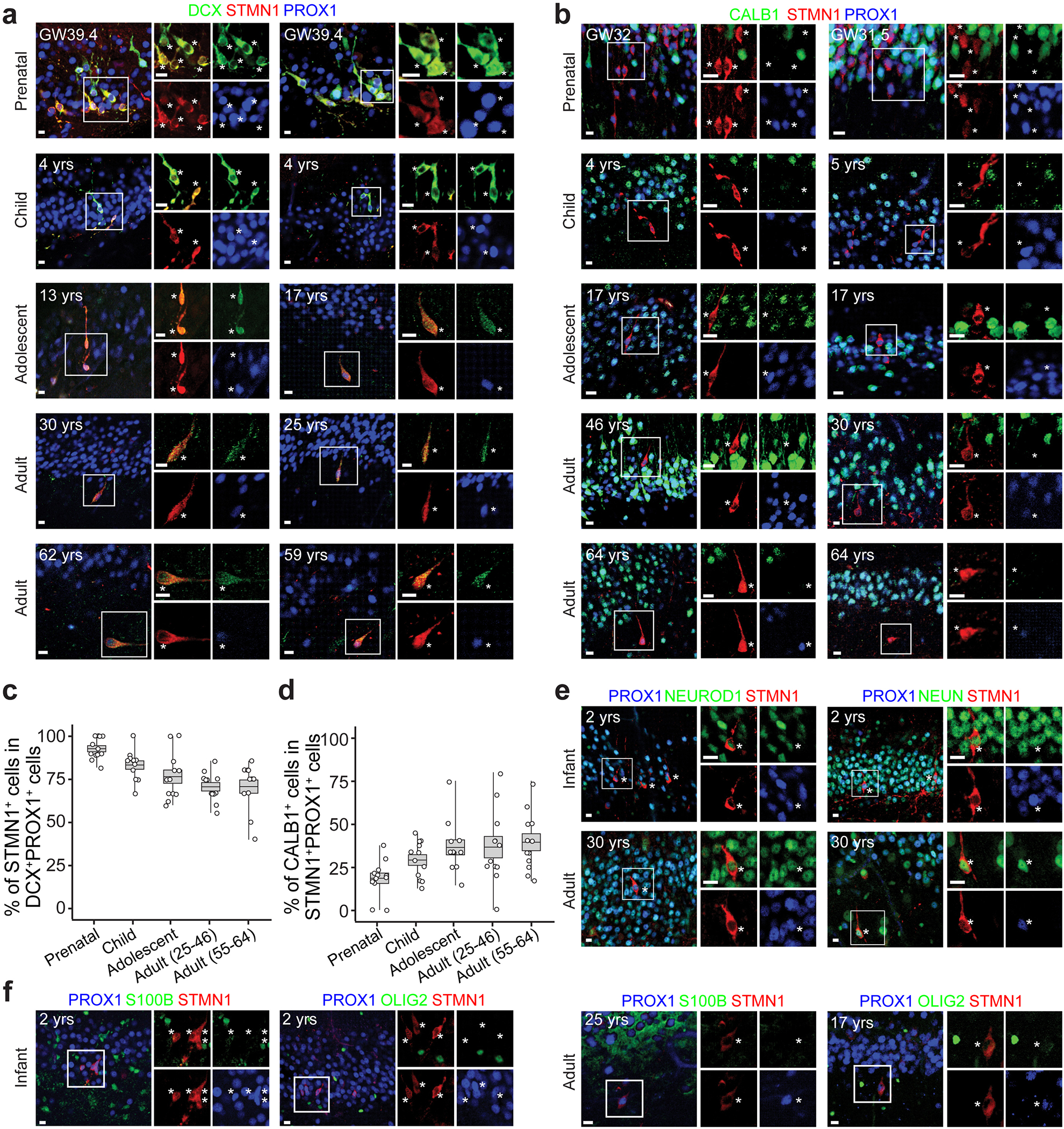
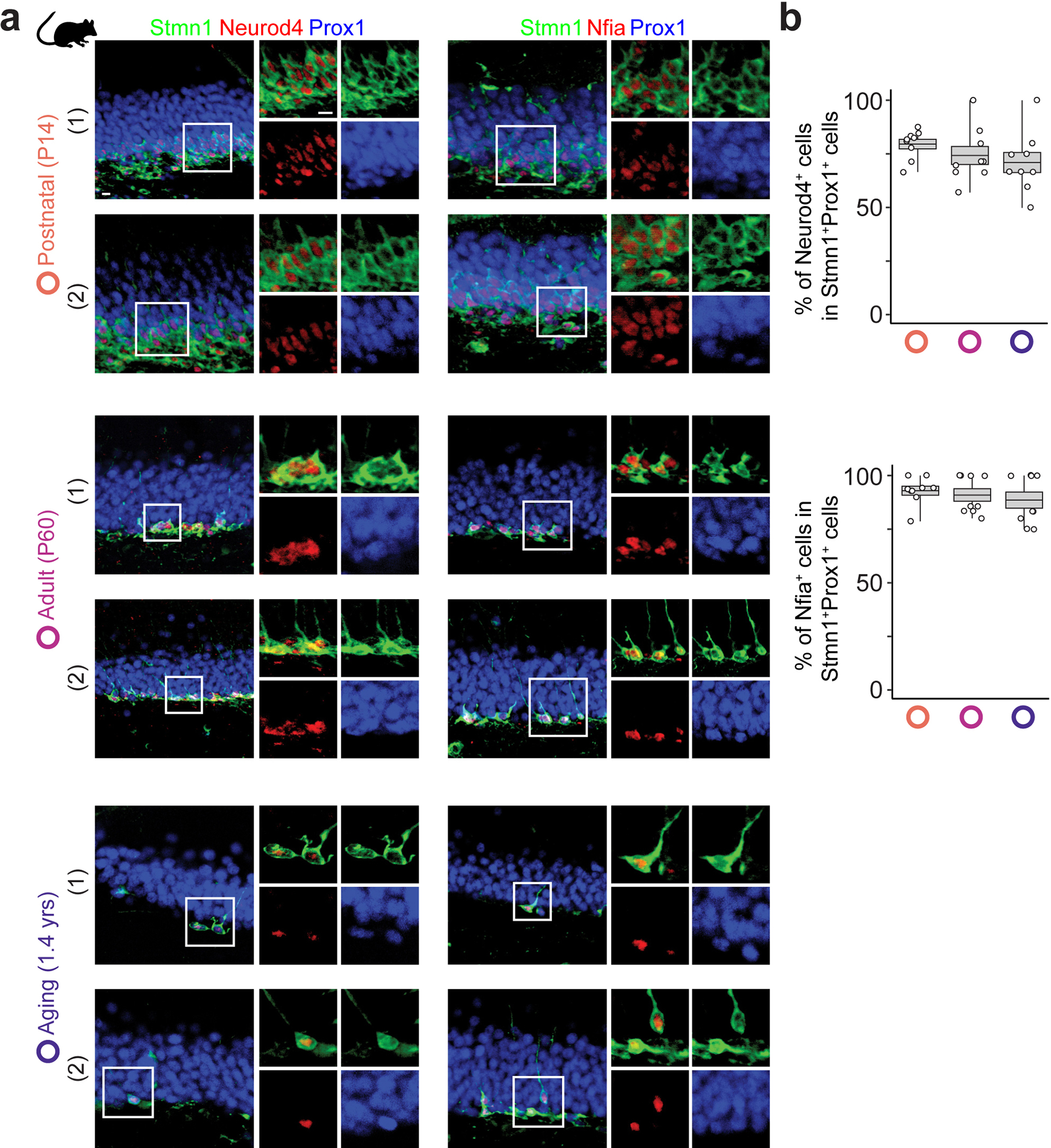
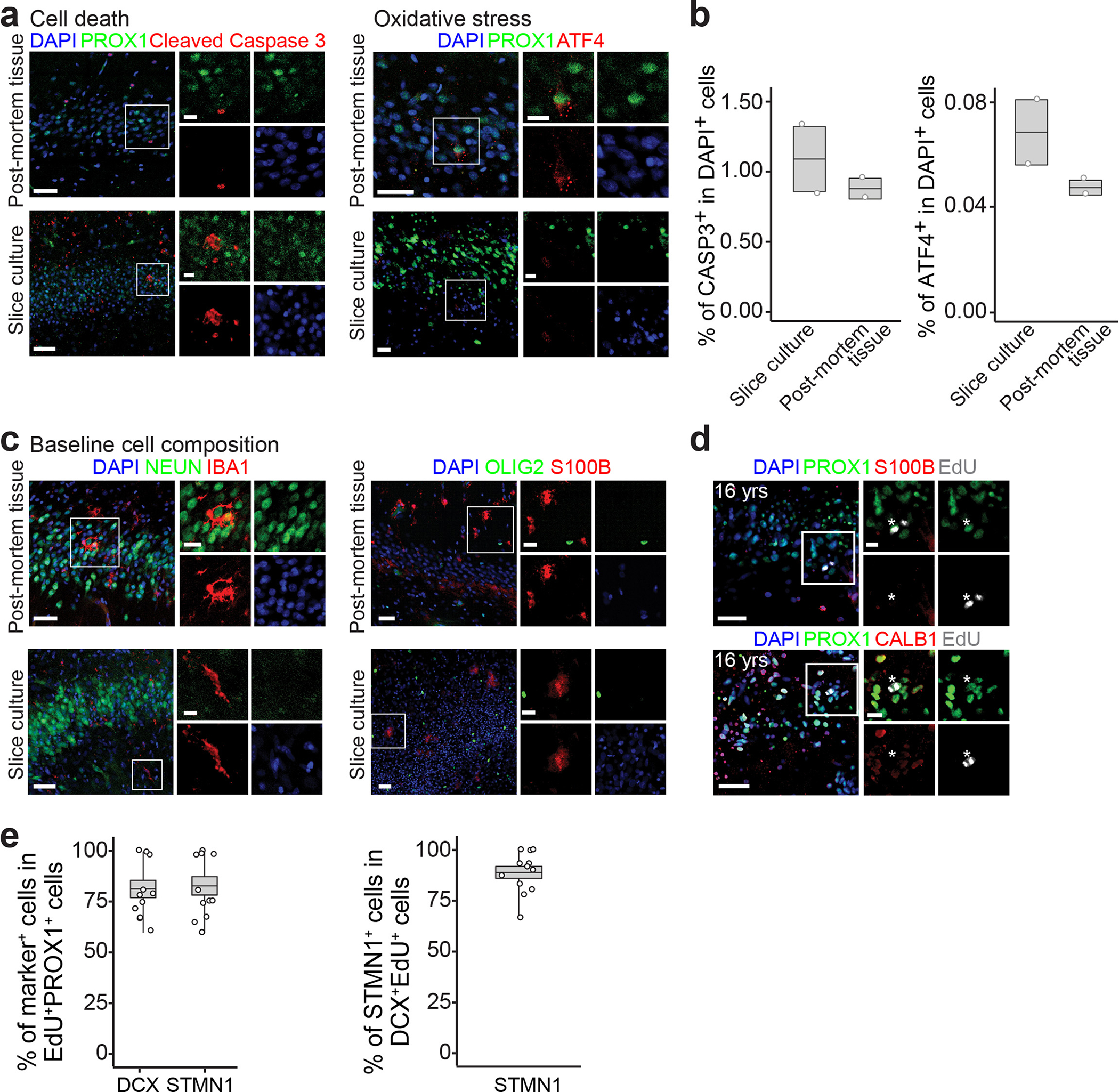
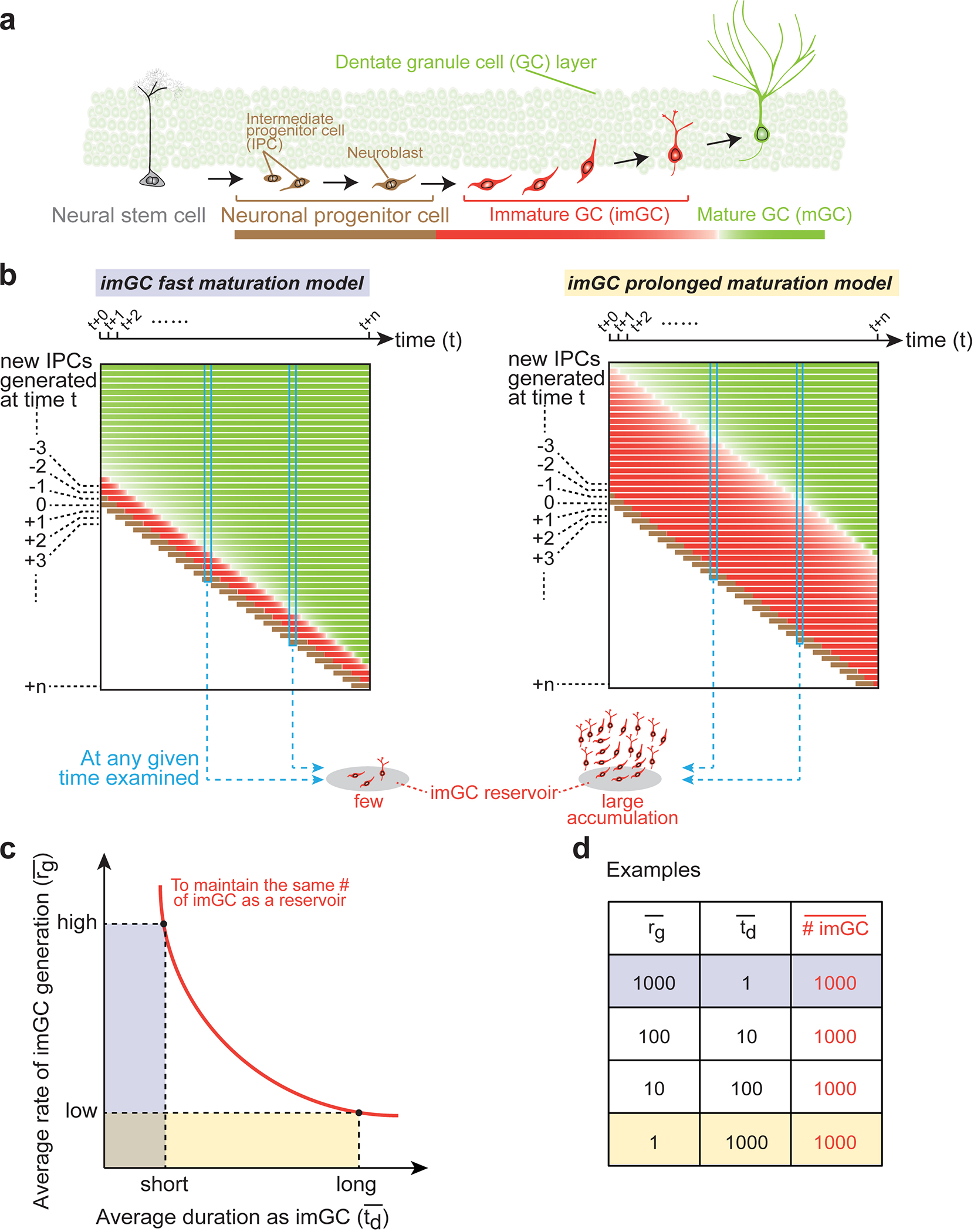
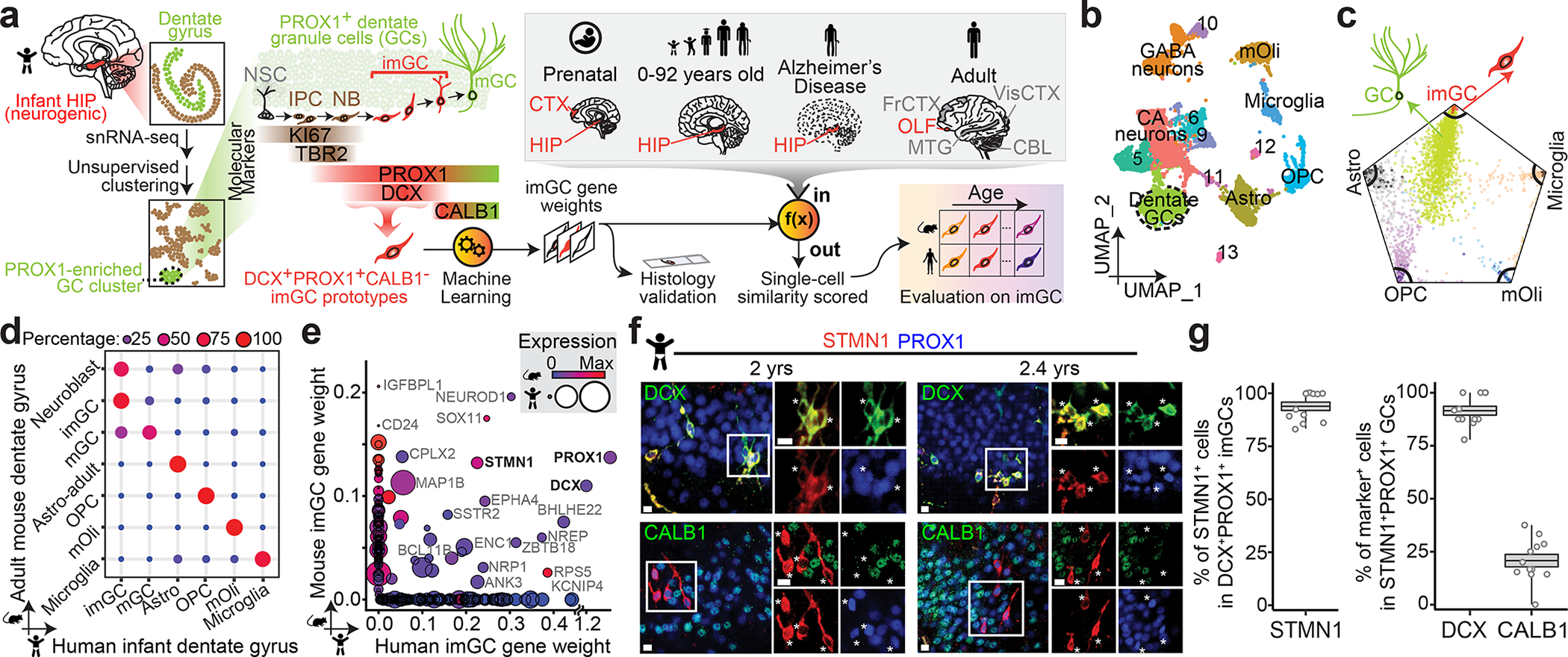
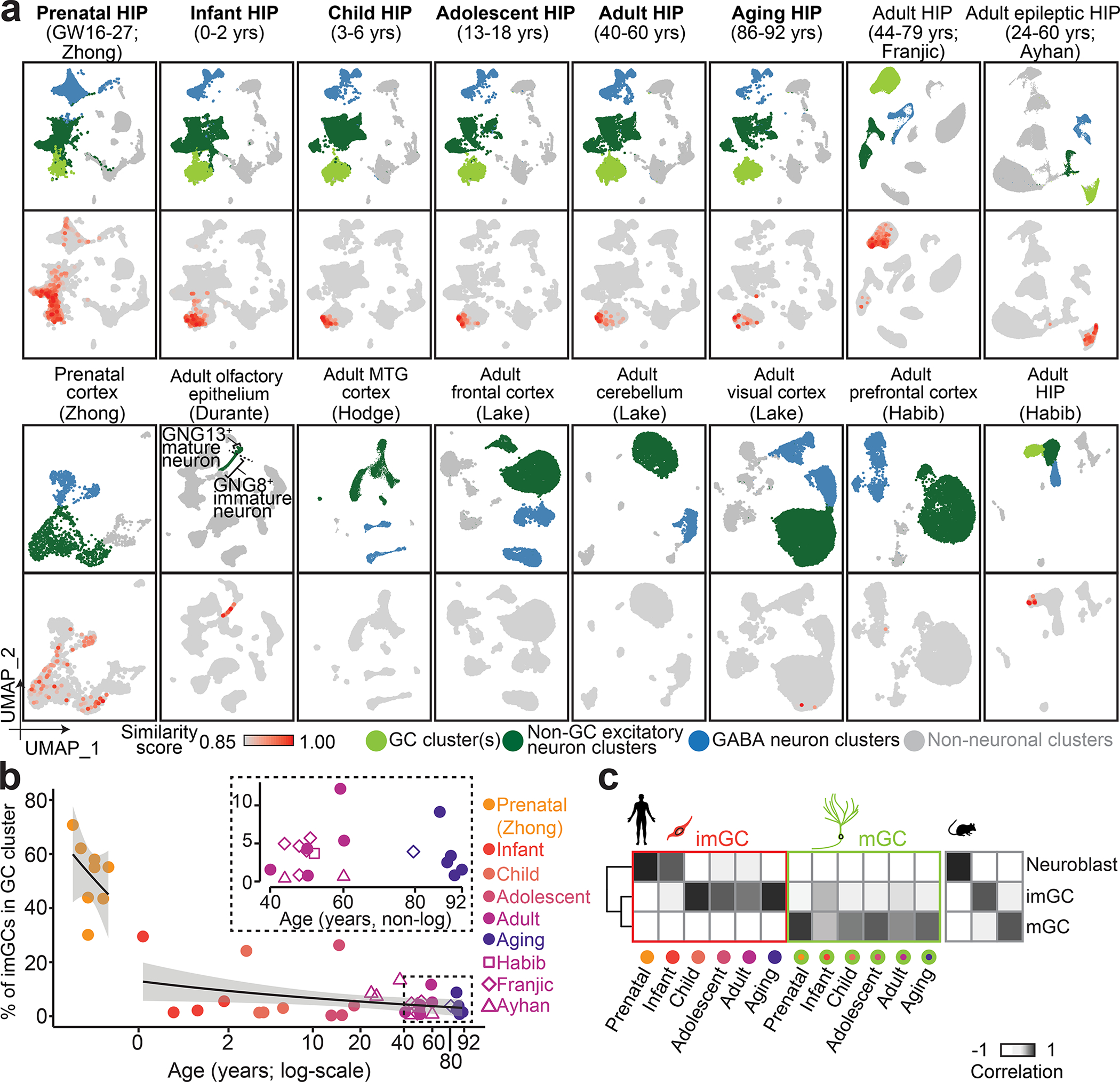
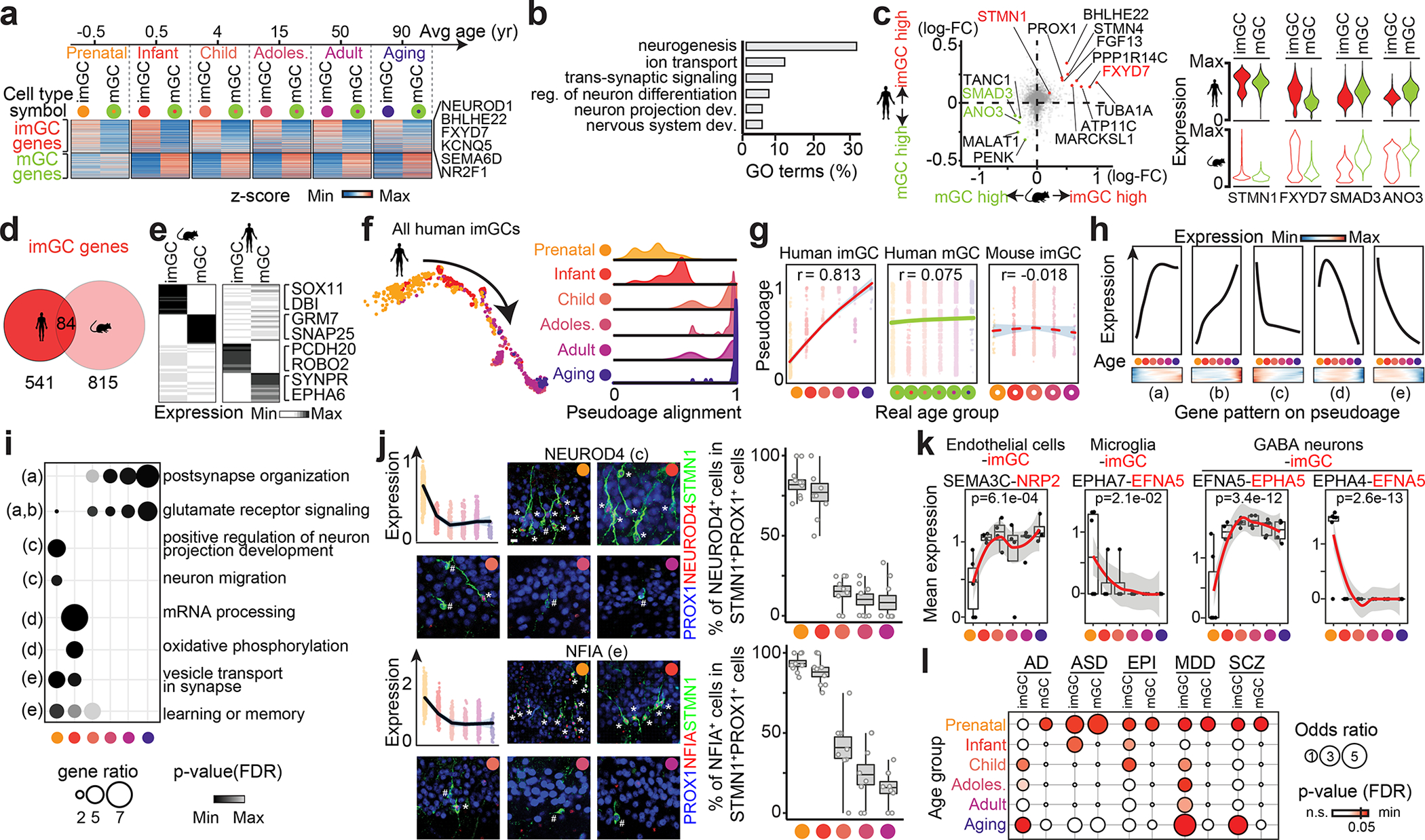
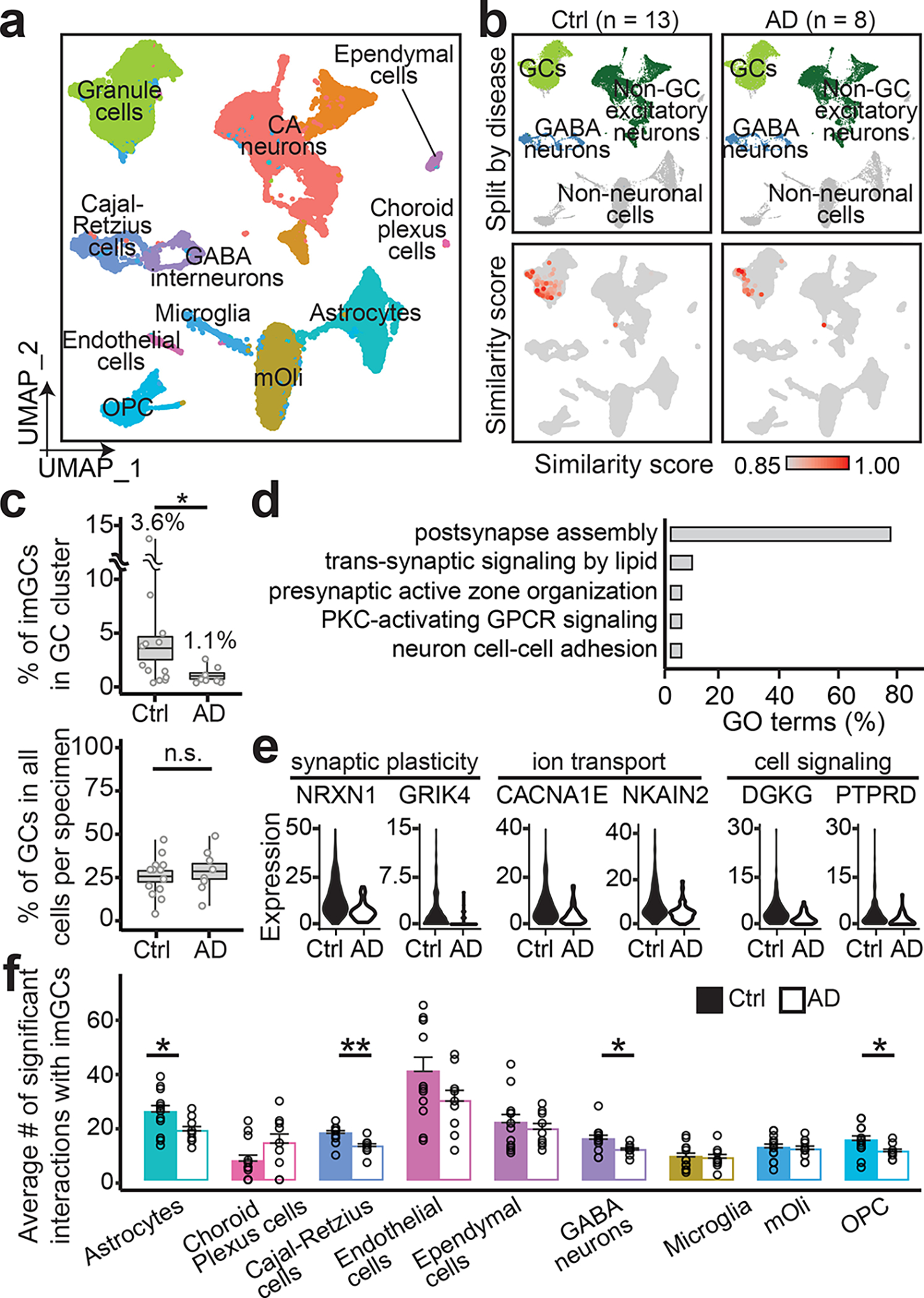
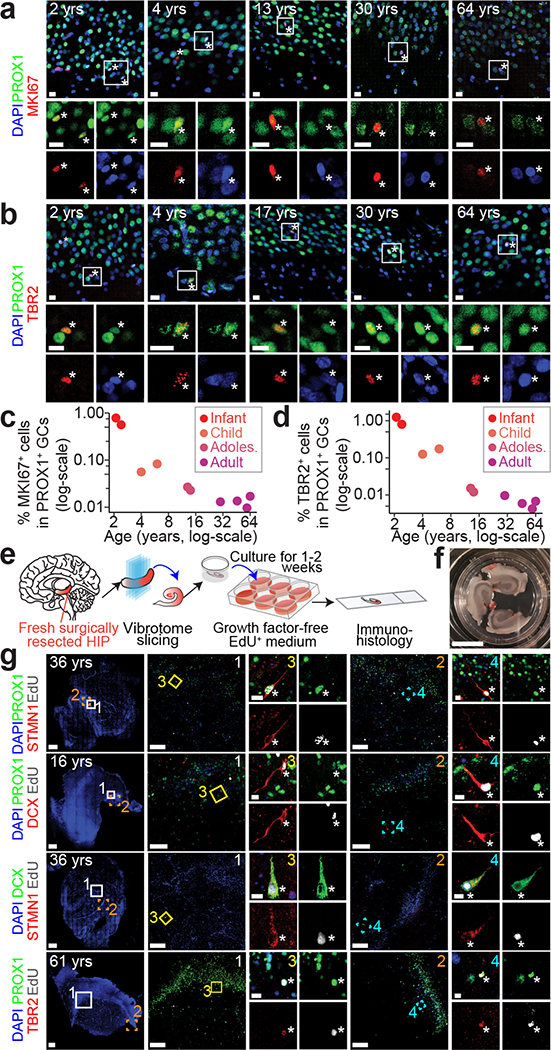
Immature dentate granule cells (imGCs) arising from adult hippocampal neurogenesis contribute to plasticity and unique brain functions in rodents1,2 and are dysregulated in multiple human neurological disorders3-5. Little is known about the molecular characteristics of adult human hippocampal imGCs, and even their existence is under debate1,6-8. Here we performed single-nucleus RNA sequencing aided by a validated machine learning-based analytic approach to identify imGCs and quantify their abundance in the human hippocampus at different stages across the lifespan. We identified common molecular hallmarks of human imGCs across the lifespan and observed age-dependent transcriptional dynamics in human imGCs that suggest changes in cellular functionality, niche interactions and disease relevance, that differ from those in mice9. We also found a decreased number of imGCs with altered gene expression in Alzheimer's disease. Finally, we demonstrated the capacity for neurogenesis in the adult human hippocampus with the presence of rare dentate granule cell fate-specific proliferating neural progenitors and with cultured surgical specimens. Together, our findings suggest the presence of a substantial number of imGCs in the adult human hippocampus via low-frequency de novo generation and protracted maturation, and our study reveals their molecular properties across the lifespan and in Alzheimer's disease.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures

References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
