

Context-specific regulation and function of mRNA alternative polyadenylation
- PMID: 35798852
- PMCID: PMC9261900
- DOI: 10.1038/s41580-022-00507-5
Context-specific regulation and function of mRNA alternative polyadenylation
Abstract
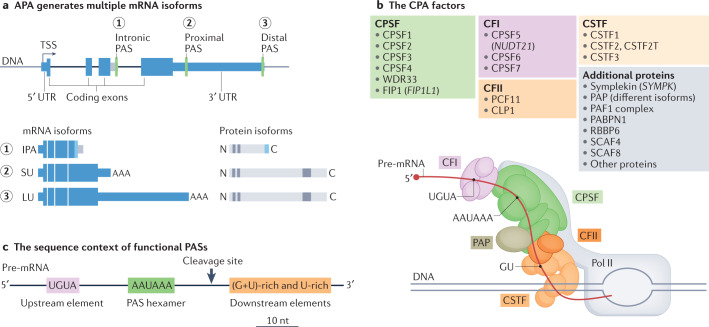
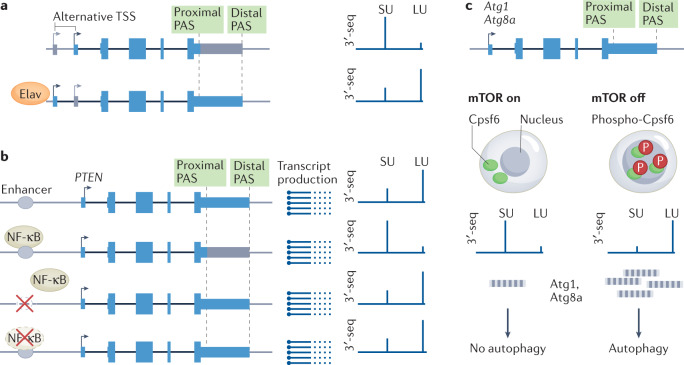
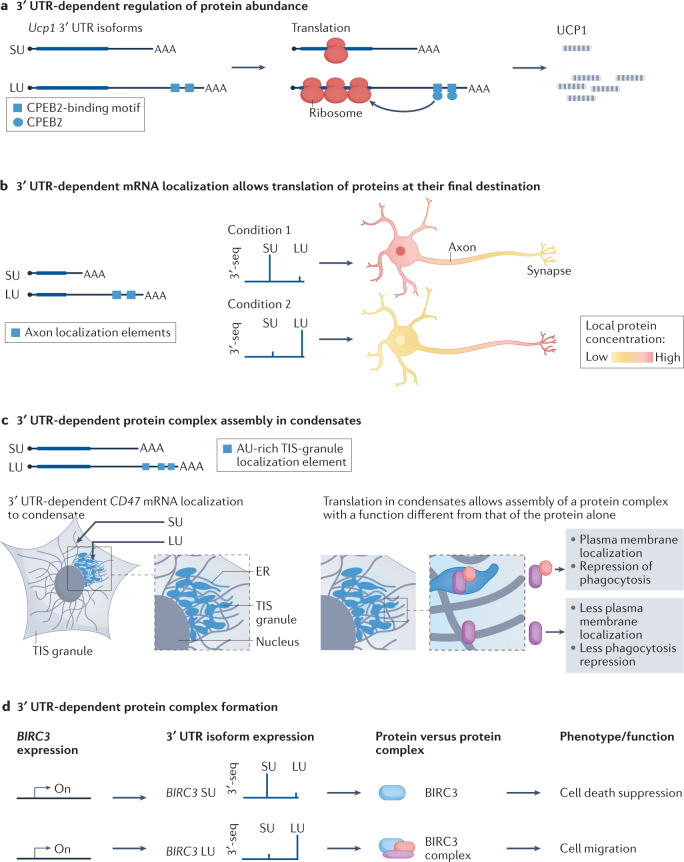
Alternative cleavage and polyadenylation (APA) is a widespread mechanism to generate mRNA isoforms with alternative 3' untranslated regions (UTRs). The expression of alternative 3' UTR isoforms is highly cell type specific and is further controlled in a gene-specific manner by environmental cues. In this Review, we discuss how the dynamic, fine-grained regulation of APA is accomplished by several mechanisms, including cis-regulatory elements in RNA and DNA and factors that control transcription, pre-mRNA cleavage and post-transcriptional processes. Furthermore, signalling pathways modulate the activity of these factors and integrate APA into gene regulatory programmes. Dysregulation of APA can reprogramme the outcome of signalling pathways and thus can control cellular responses to environmental changes. In addition to the regulation of protein abundance, APA has emerged as a major regulator of mRNA localization and the spatial organization of protein synthesis. This role enables the regulation of protein function through the addition of post-translational modifications or the formation of protein-protein interactions. We further discuss recent transformative advances in single-cell RNA sequencing and CRISPR-Cas technologies, which enable the mapping and functional characterization of alternative 3' UTRs in any biological context. Finally, we discuss new APA-based RNA therapeutics, including compounds that target APA in cancer and therapeutic genome editing of degenerative diseases.
© 2022. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Fansler MM, Zhen G, Mayr C. Quantification of alternative 3′UTR isoforms from single cell RNA-seq data with scUTRquant. bioRxiv. 2021 doi: 10.1101/2021.11.22.469635. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
