

Protein palmitoylation-mediated palmitic acid sensing causes blood-testis barrier damage via inducing ER stress
- PMID: 35803125
- PMCID: PMC9287734
- DOI: 10.1016/j.redox.2022.102380
Protein palmitoylation-mediated palmitic acid sensing causes blood-testis barrier damage via inducing ER stress
Abstract
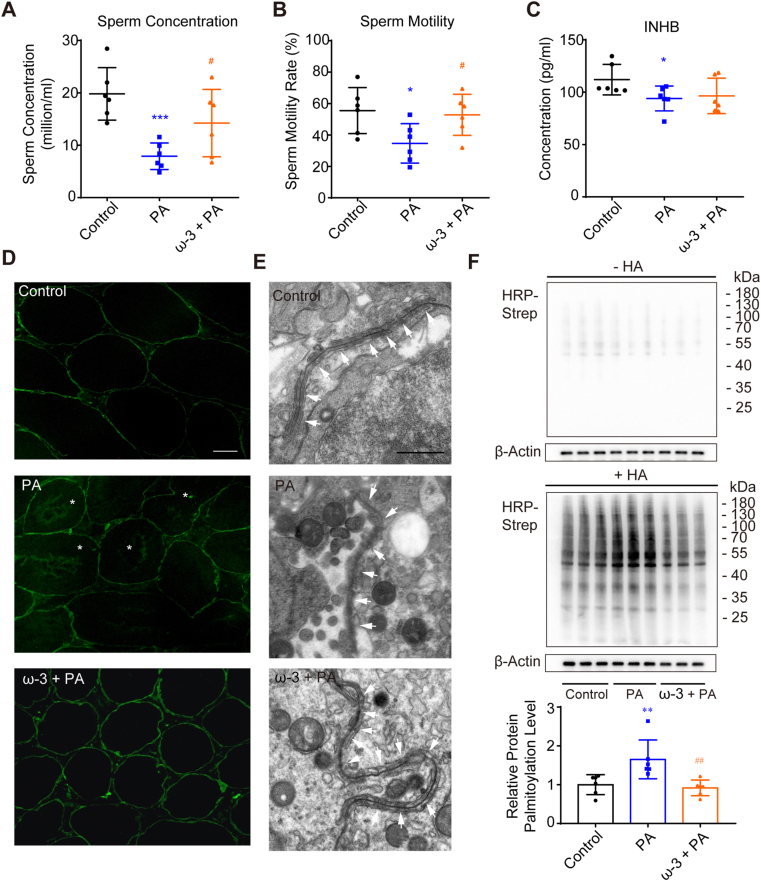
Blood-testis barrier (BTB) damage promotes spermatogenesis dysfunction, which is a critical cause of male infertility. Dyslipidemia has been correlated with male infertility, but the major hazardous lipid and the underlying mechanism remains unclear. In this study, we firstly discovered an elevation of palmitic acid (PA) and a decrease of inhibin B in patients with severe dyszoospermia, which leaded us to explore the effects of PA on Sertoli cells. We observed a damage of BTB by PA. PA penetration to endoplasmic reticulum (ER) and its damage to ER structures were exhibited by microimaging and dynamic observation, and consequent ER stress was proved to mediate PA-induced Sertoli cell barrier disruption. Remarkably, we demonstrated a critical role of aberrant protein palmitoylation in PA-induced Sertoli cell barrier dysfunction. An ER protein, Calnexin, was screened out and was demonstrated to participate in this process, and suppression of its palmitoylation showed an ameliorating effect. We also found that ω-3 poly-unsaturated fatty acids down-regulated Calnexin palmitoylation, and alleviated BTB dysfunction. Our results indicate that dysregulated palmitoylation induced by PA plays a pivotal role in BTB disruption and subsequent spermatogenesis dysfunction, suggesting that protein palmitoylation might be therapeutically targetable in male infertility.
Keywords: Blood-testis barrier; Endoplasmic reticulum stress; Palmitic acid; Palmitoylation; Sertoli cell; Tight junction.
Copyright © 2022 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures









References
-
- McCabe M.J., Tarulli G.A., Laven-Law G., Matthiesson K.L., Meachem S.J., McLachlan R.I., Dinger M.E., Stanton P.G. Gonadotropin suppression in men leads to a reduction in claudin-11 at the Sertoli cell tight junction. Hum. Reprod. 2016;31(4):875–886. [pii] - PubMed
-
- Zhu R., Wang J., Feng T., Hu X., Jiang C., Wang X., Li K., Sang Y., Hua Y., Sun H., Yao B., Li C. The alteration of RhoA geranylgeranylation and Ras farnesylation breaks the integrity of the blood-testis barrier and results in hypospermatogenesis. Cell Death Dis. 2019;10(6):450. doi: 10.1038/s41419-019-1688-9. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
