

Theory of the Multiregional Neocortex: Large-Scale Neural Dynamics and Distributed Cognition
- PMID: 35803587
- PMCID: PMC11583996
- DOI: 10.1146/annurev-neuro-110920-035434
Theory of the Multiregional Neocortex: Large-Scale Neural Dynamics and Distributed Cognition
Abstract
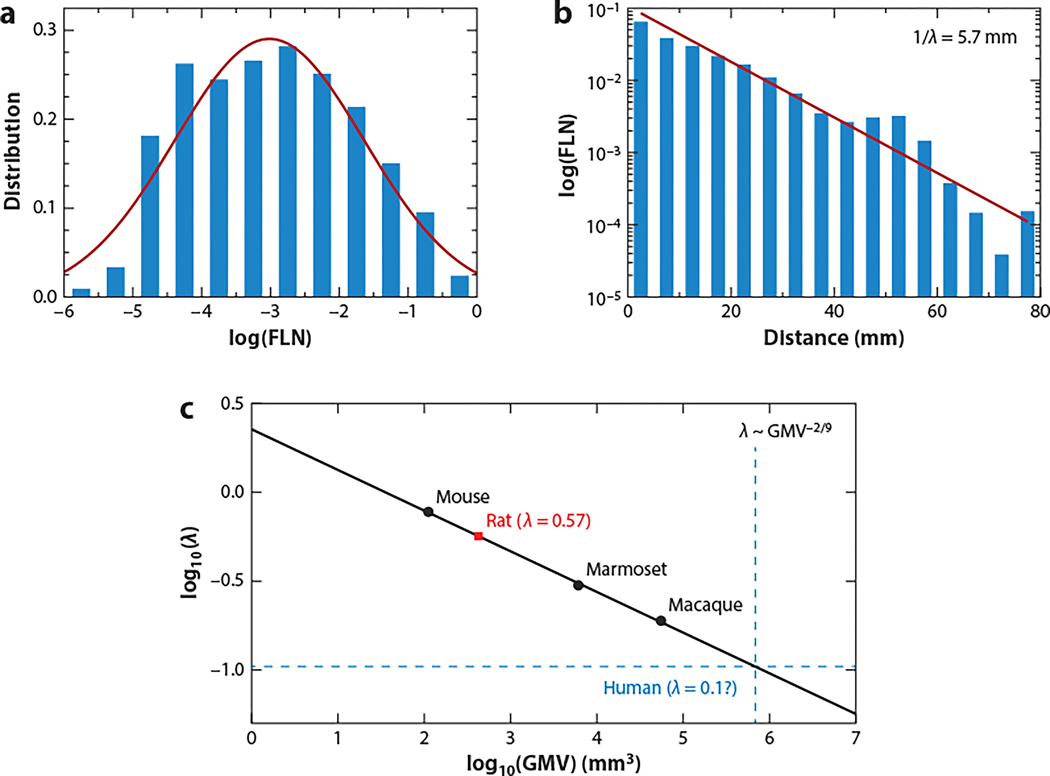
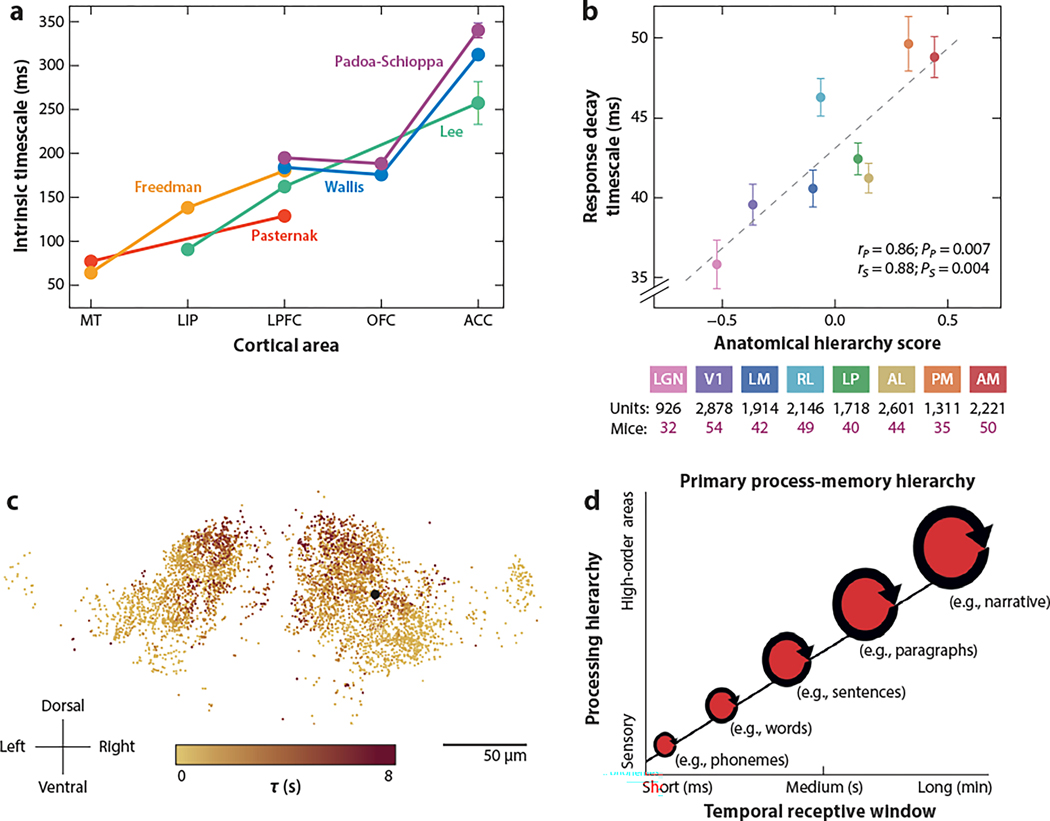
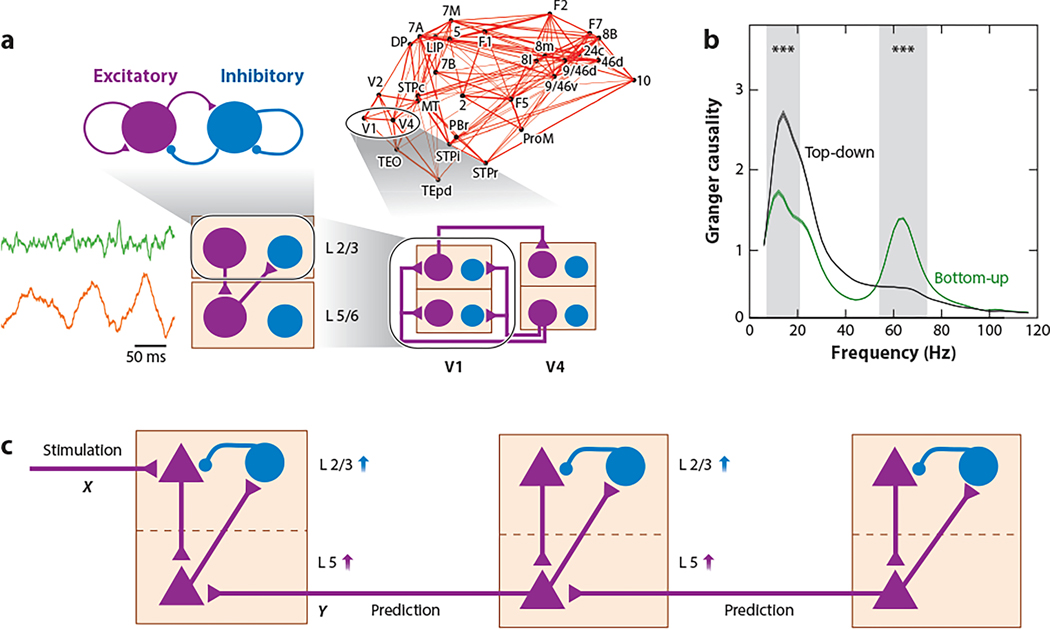
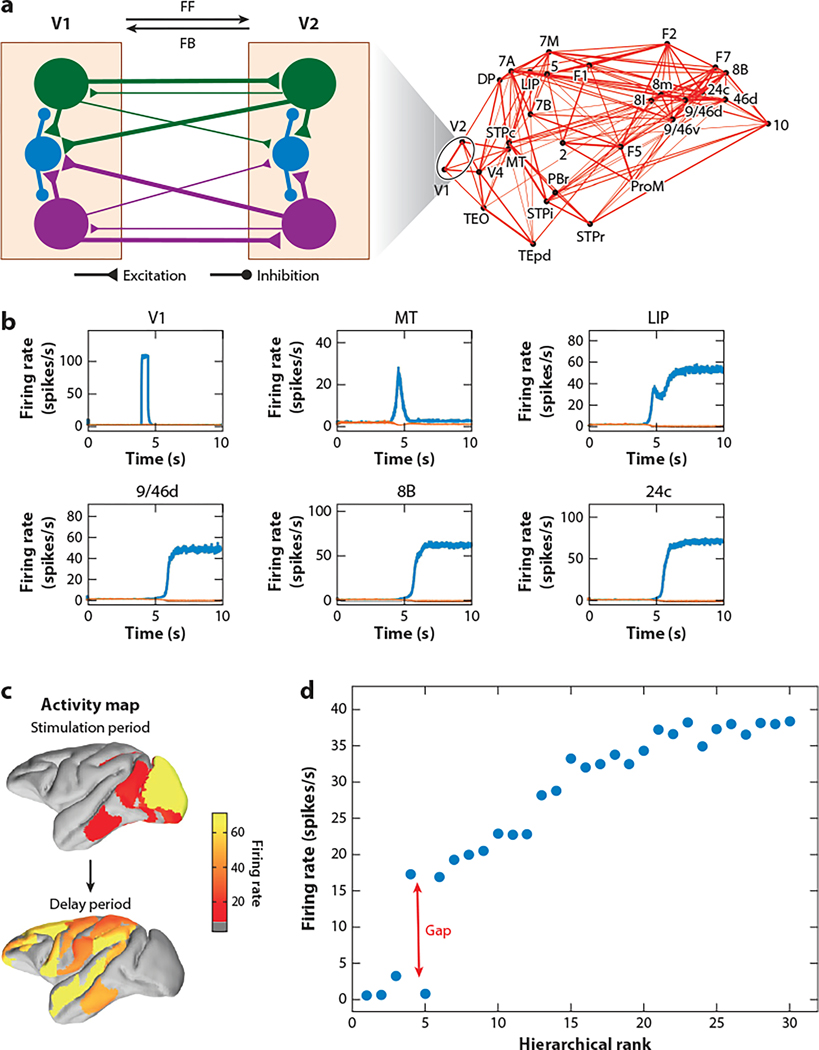
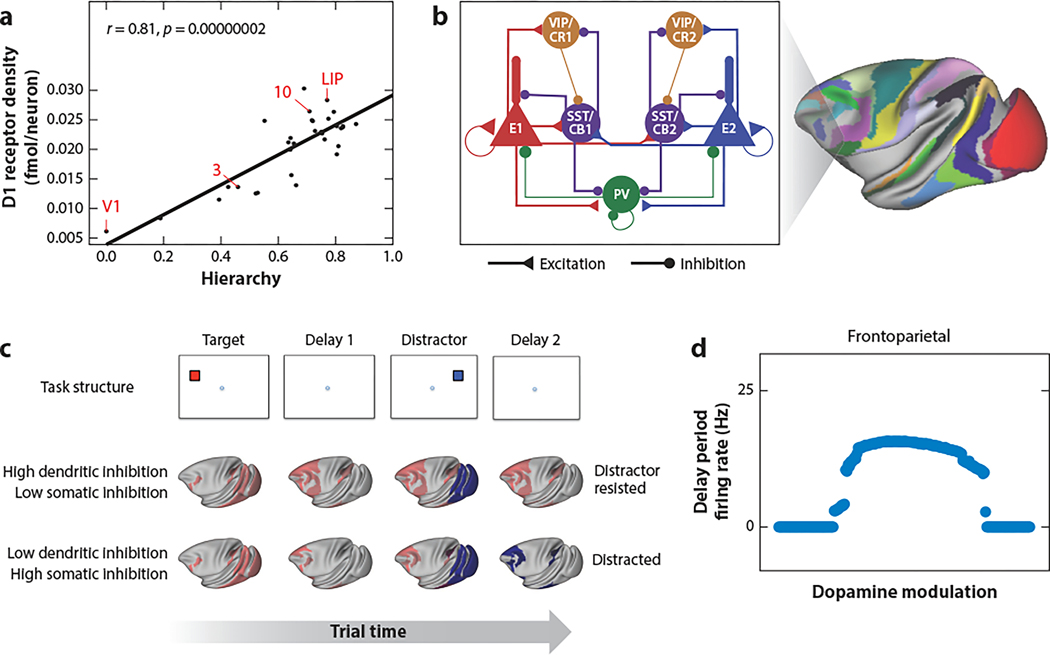
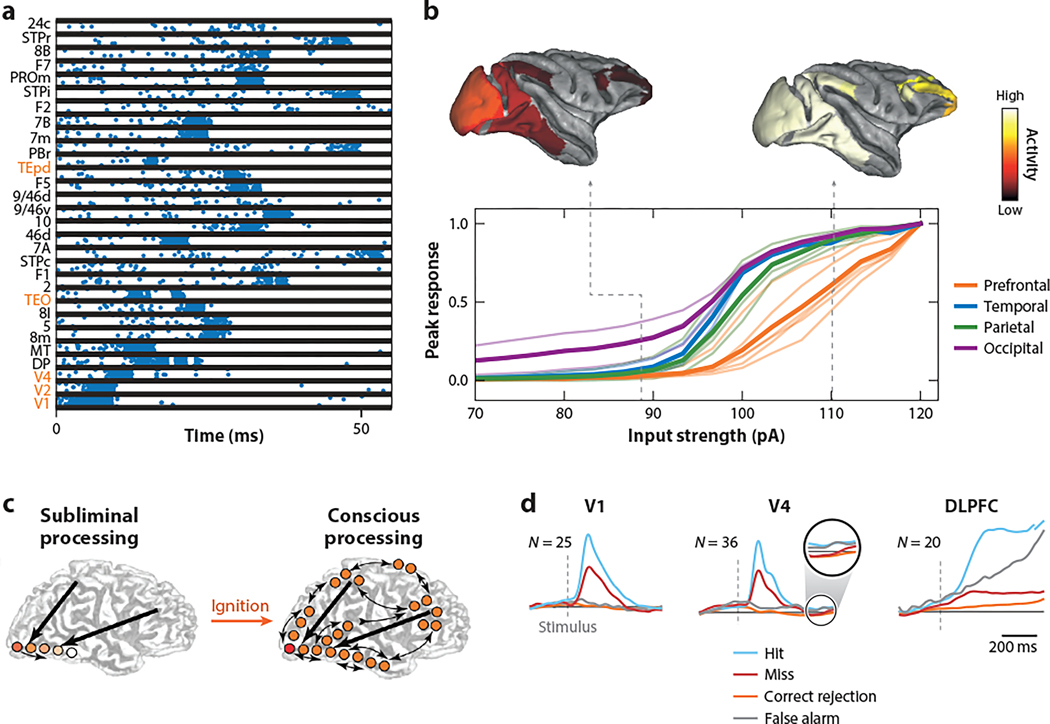
The neocortex is a complex neurobiological system with many interacting regions. How these regions work together to subserve flexible behavior and cognition has become increasingly amenable to rigorous research. Here, I review recent experimental and theoretical work on the modus operandi of a multiregional cortex. These studies revealed several general principles for the neocortical interareal connectivity, low-dimensional macroscopic gradients of biological properties across cortical areas, and a hierarchy of timescales for information processing. Theoretical work suggests testable predictions regarding differential excitation and inhibition along feedforward and feedback pathways in the cortical hierarchy. Furthermore, modeling of distributed working memory and simple decision-making has given rise to a novel mathematical concept, dubbed bifurcation in space, that potentially explains how different cortical areas, with a canonical circuit organization but gradients of biological heterogeneities, are able to subserve their respective (e.g., sensory coding versus executive control) functions in a modularly organized brain.
Keywords: computational modeling; distributed cognition; global brain dynamics; hierarchy of timescales; macroscopic gradients; neocortical connectome.
Figures
References
-
- Abbott LF, Bock DD, Callaway EM, Denk W, Dulac C, et al. 2020. The mind of a mouse. Cell 182:1372–76 - PubMed
-
- Amit D. 1995. The Hebbian paradigm reintegrated: local reverberations as internal representations. Behav. Brain Sci 18:617–25
-
- Amunts K, Zilles K. 2015. Architectonic mapping of the human brain beyond Brodmann. Neuron 88:1086–107 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
