

Morphogenetic Roles of Hydrostatic Pressure in Animal Development
- PMID: 35804476
- PMCID: PMC9675319
- DOI: 10.1146/annurev-cellbio-120320-033250
Morphogenetic Roles of Hydrostatic Pressure in Animal Development
Abstract
During organismal development, organs and systems are built following a genetic blueprint that produces structures capable of performing specific physiological functions. Interestingly, we have learned that the physiological activities of developing tissues also contribute to their own morphogenesis. Specifically, physiological activities such as fluid secretion and cell contractility generate hydrostatic pressure that can act as a morphogenetic force. Here, we first review the role of hydrostatic pressure in tube formation during animal development and discuss mathematical models of lumen formation. We then illustrate specific roles of the notochord as a hydrostatic scaffold in anterior-posterior axis development in chordates. Finally, we cover some examples of how fluid flows influence morphogenetic processes in other developmental contexts. Understanding how fluid forces act during development will be key for uncovering the self-organizing principles that control morphogenesis.
Keywords: axis elongation; hydrostatic pressure; lumen; morphogenesis; notochord; tube formation.
Figures
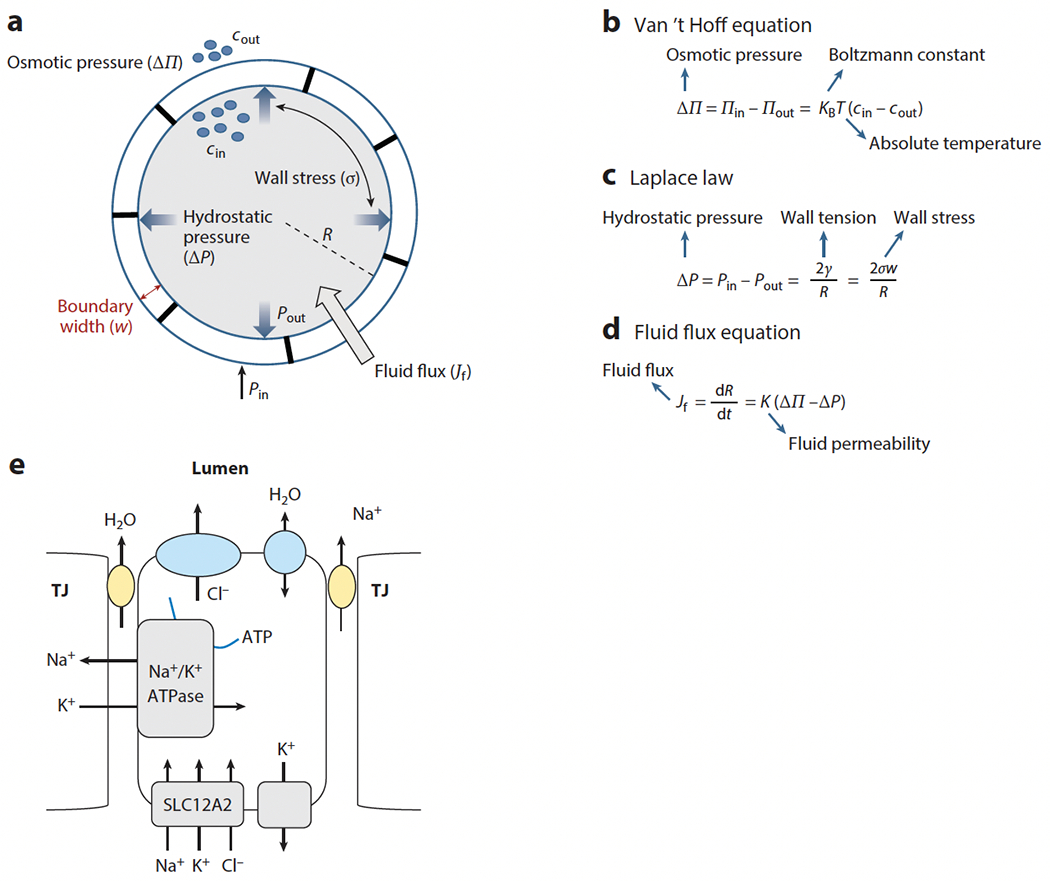
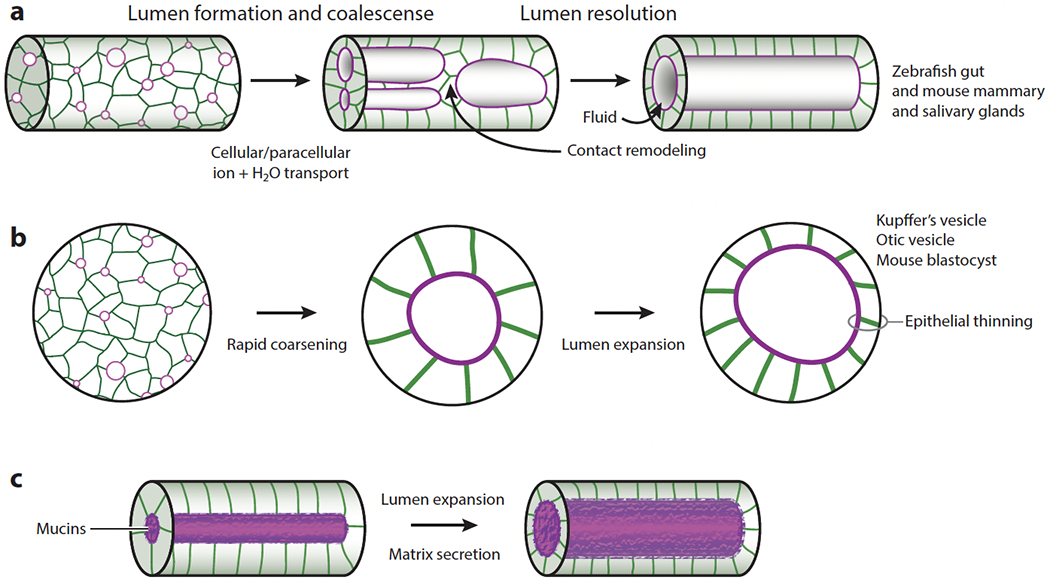
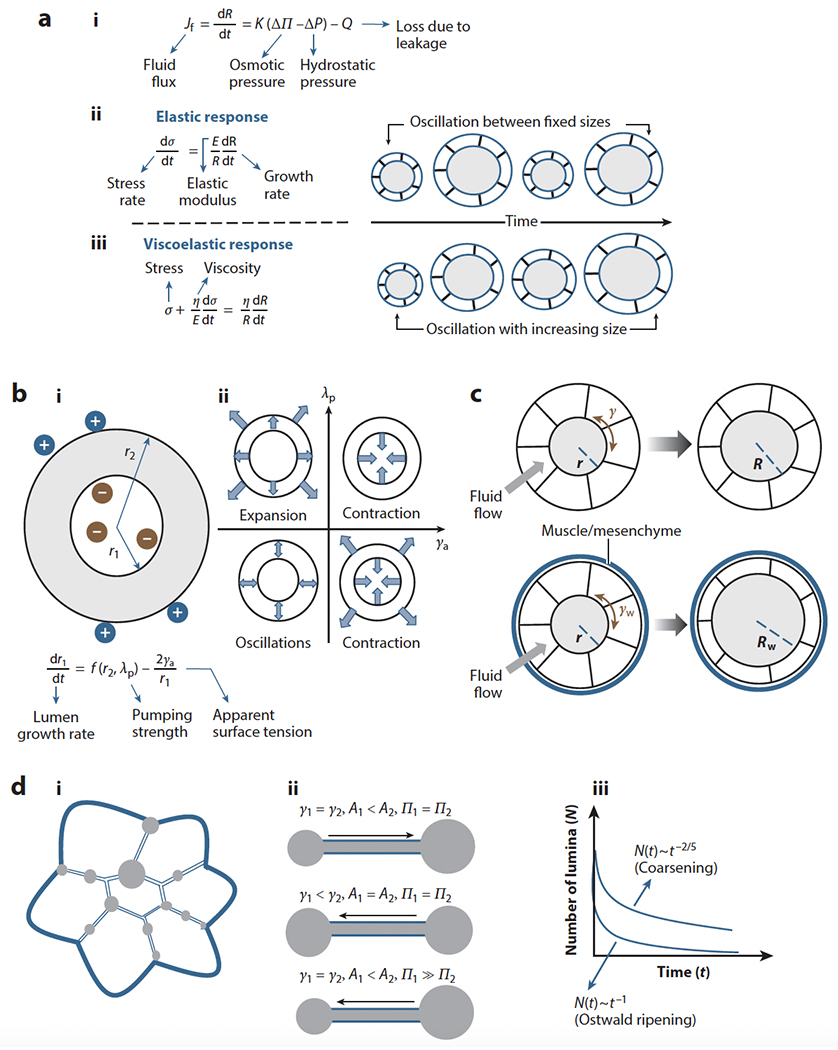
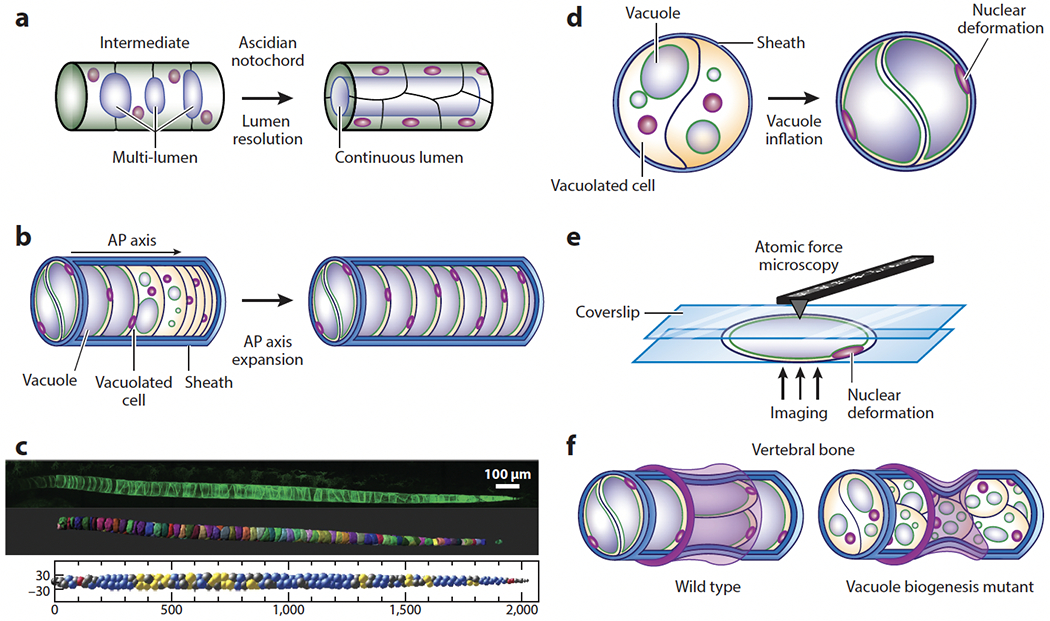
References
-
- Adams DS, Keller R, Koehl MA. 1990. The mechanics of notochord elongation, straightening and stiffening in the embryo of Xenopus laevis. Development 110:115–30 - PubMed
-
- Bagnat M, Cheung ID, Mostov KE, Stainier DY. 2007. Genetic control of single lumen formation in the zebrafish gut. Nat. Cell Biol 9:954–60 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
