

Extracellular Cysteines Are Critical to Form Functional Cx46 Hemichannels
- PMID: 35806258
- PMCID: PMC9266770
- DOI: 10.3390/ijms23137252
Extracellular Cysteines Are Critical to Form Functional Cx46 Hemichannels
Abstract
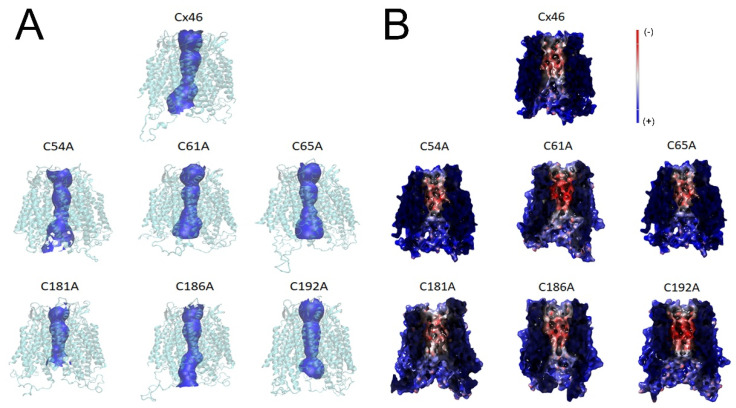
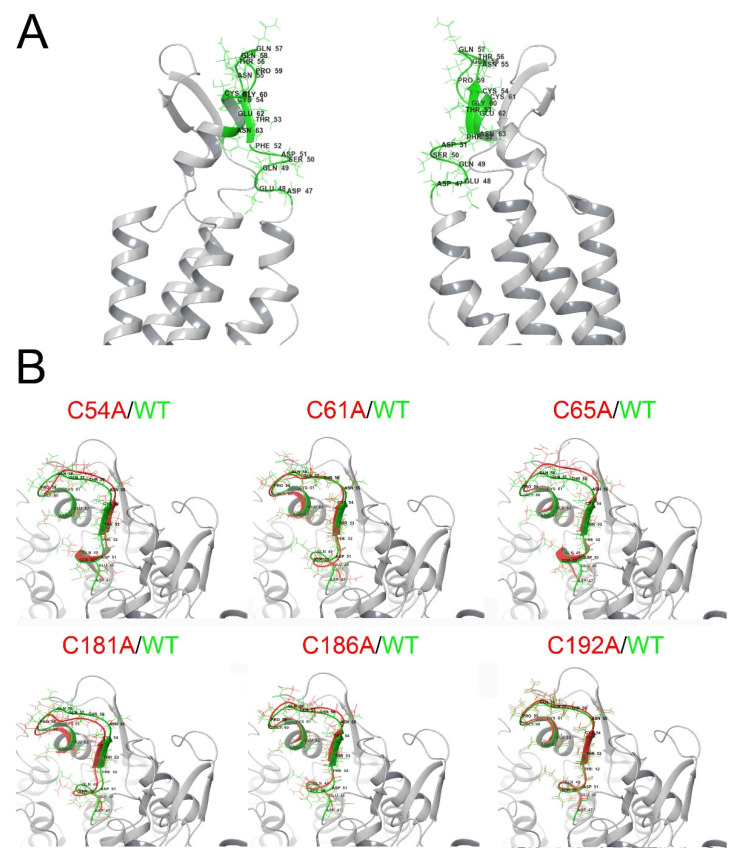
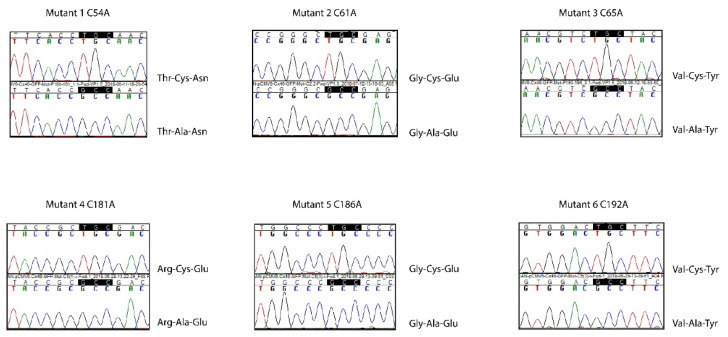
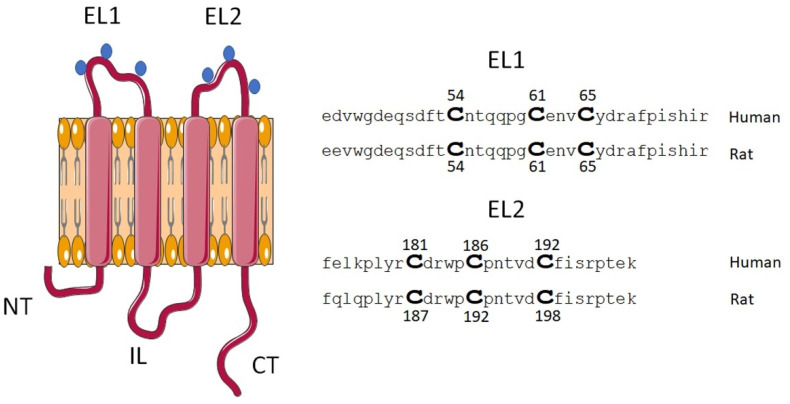
Connexin (Cxs) hemichannels participate in several physiological and pathological processes, but the molecular mechanisms that control their gating remain elusive. We aimed at determining the role of extracellular cysteines (Cys) in the gating and function of Cx46 hemichannels. We studied Cx46 and mutated all of its extracellular Cys to alanine (Ala) (one at a time) and studied the effects of the Cys mutations on Cx46 expression, localization, and hemichannel activity. Wild-type Cx46 and Cys mutants were expressed at comparable levels, with similar cellular localization. However, functional experiments showed that hemichannels formed by the Cys mutants did not open either in response to membrane depolarization or removal of extracellular divalent cations. Molecular-dynamics simulations showed that Cys mutants may show a possible alteration in the electrostatic potential of the hemichannel pore and an altered disposition of important residues that could contribute to the selectivity and voltage dependency in the hemichannels. Replacement of extracellular Cys resulted in "permanently closed hemichannels", which is congruent with the inhibition of the Cx46 hemichannel by lipid peroxides, through the oxidation of extracellular Cys. These results point to the modification of extracellular Cys as potential targets for the treatment of Cx46-hemichannel associated pathologies, such as cataracts and cancer, and may shed light into the gating mechanisms of other Cx hemichannels.
Keywords: channel permeability; connexins; extracellular loops; post-translational modification; redox sensing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
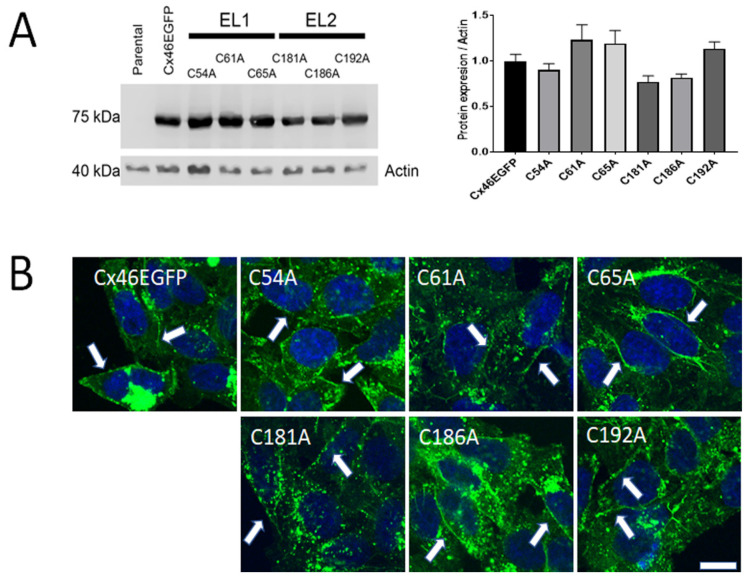
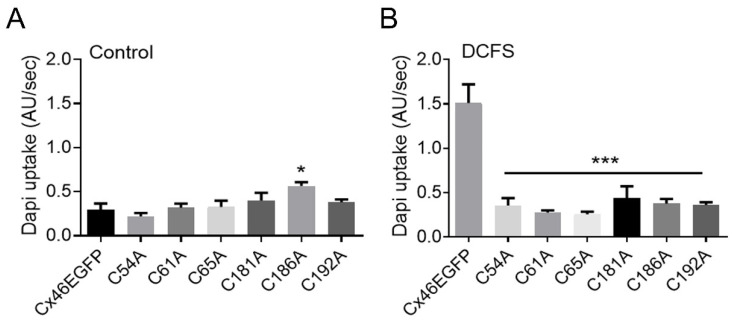
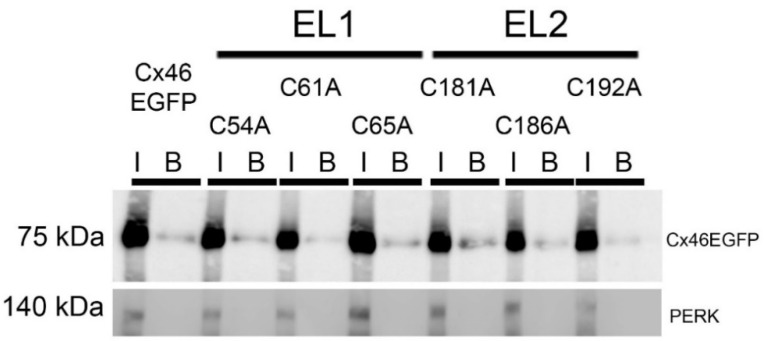
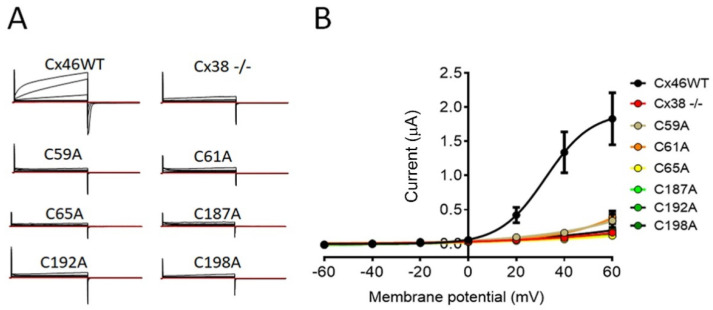
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
