

The Anti-Inflammatory Properties of Chaga Extracts Obtained by Different Extraction Methods against LPS-Induced RAW 264.7
- PMID: 35807453
- PMCID: PMC9268247
- DOI: 10.3390/molecules27134207
The Anti-Inflammatory Properties of Chaga Extracts Obtained by Different Extraction Methods against LPS-Induced RAW 264.7
Abstract
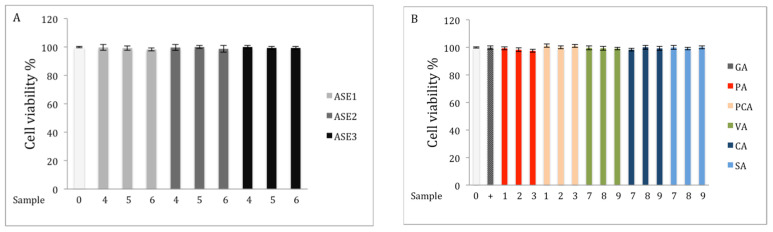
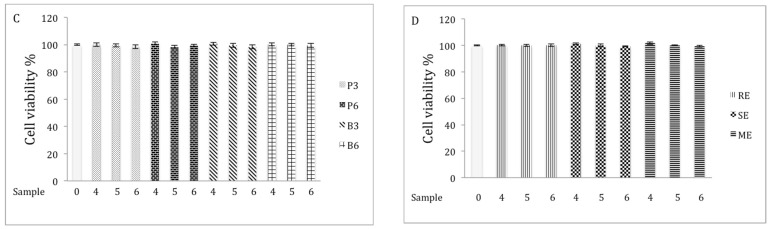
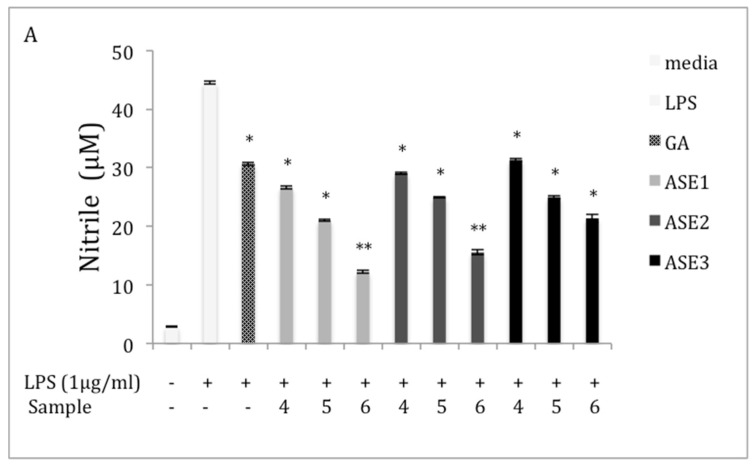
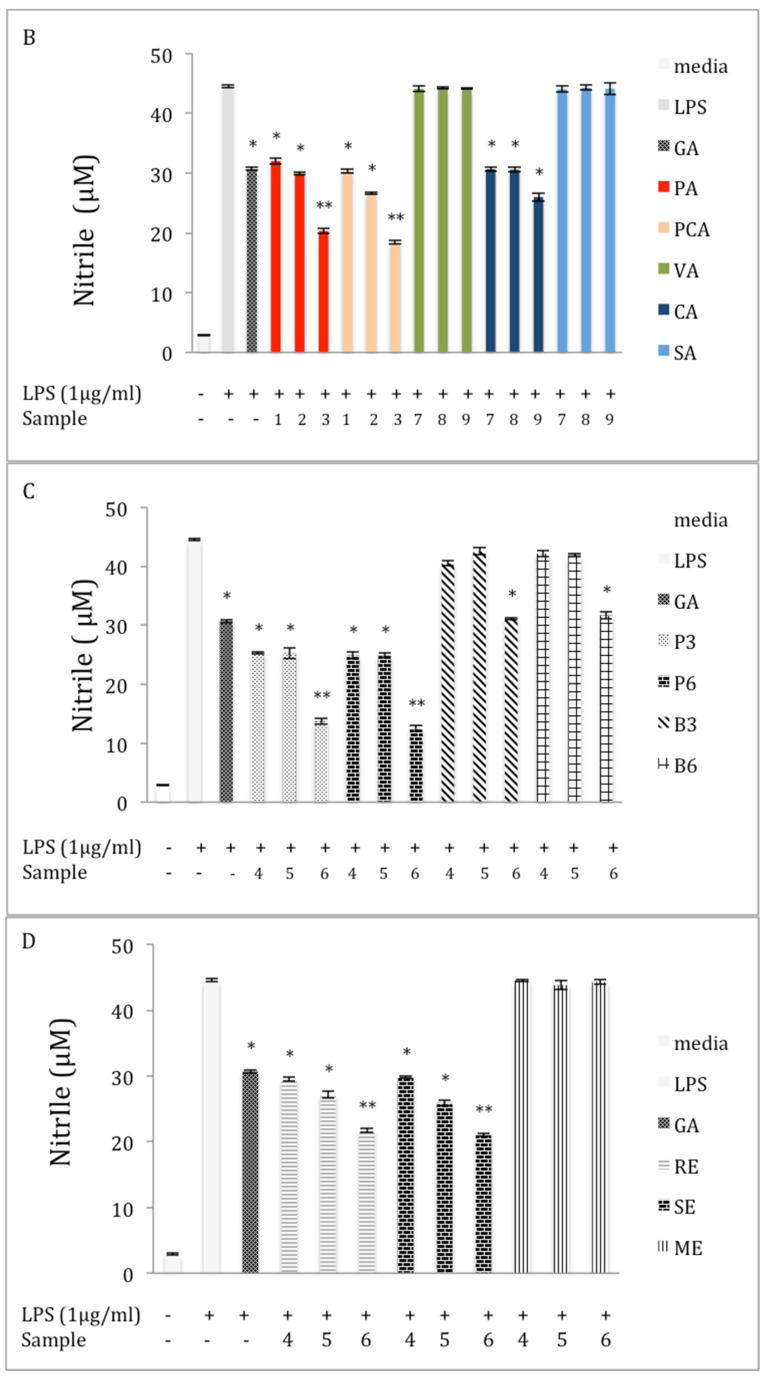
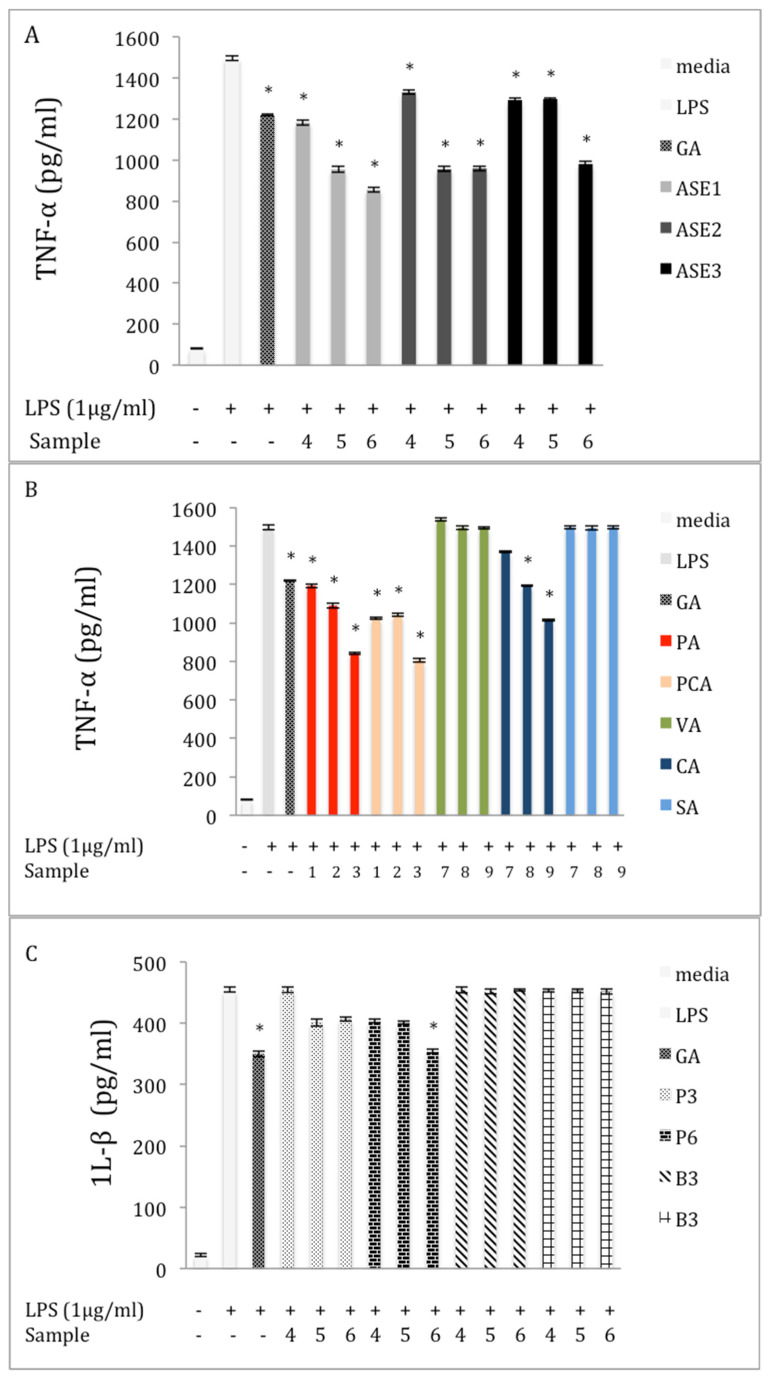
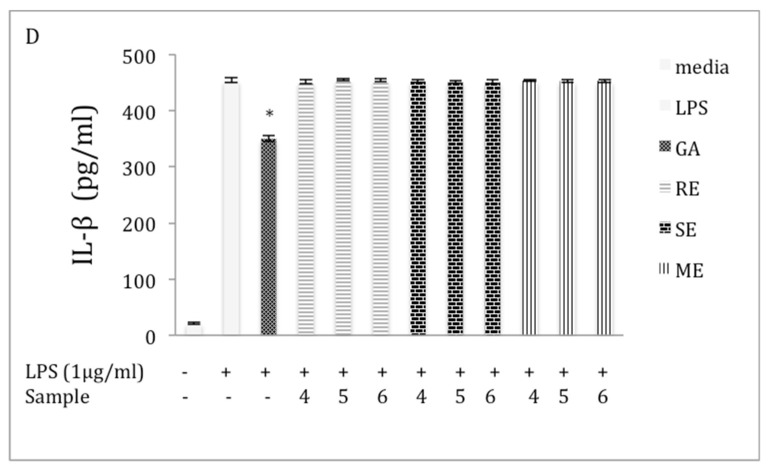
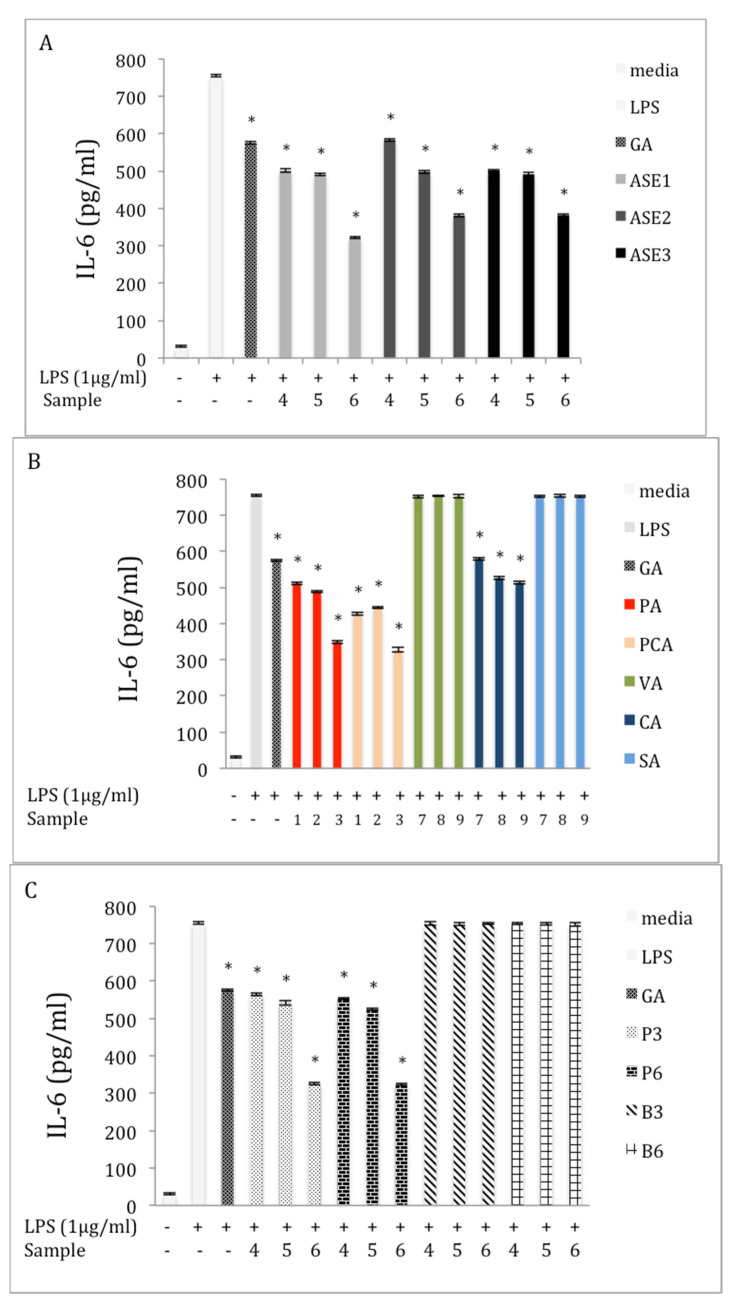
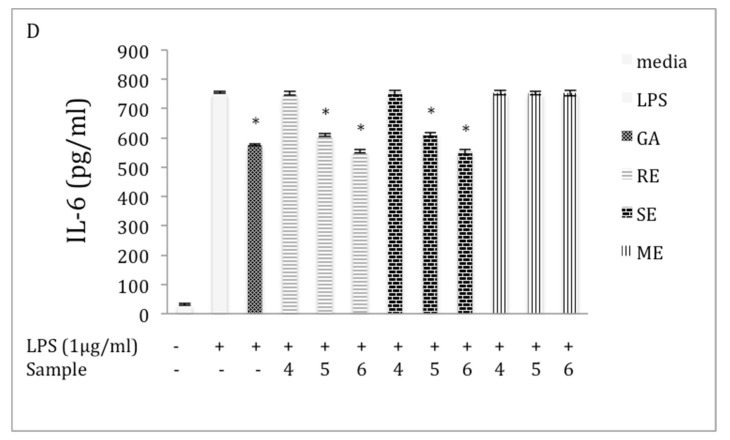
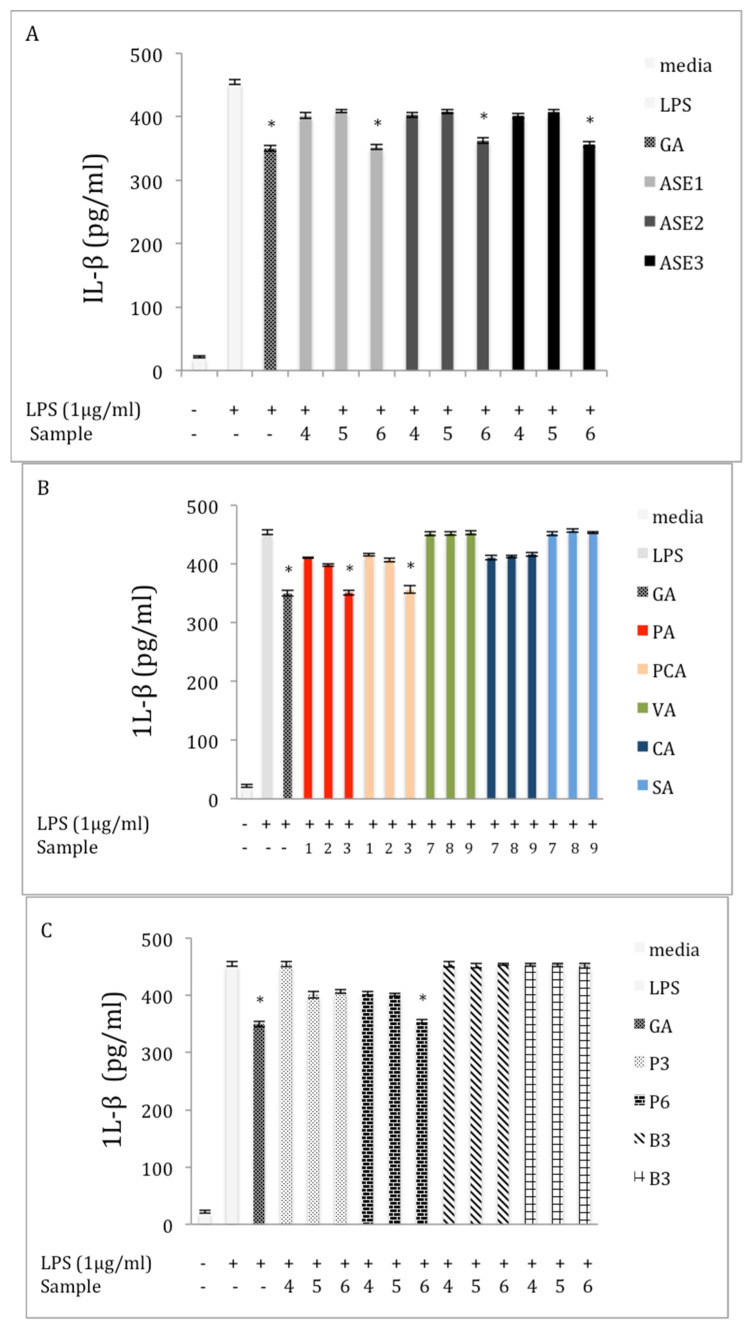
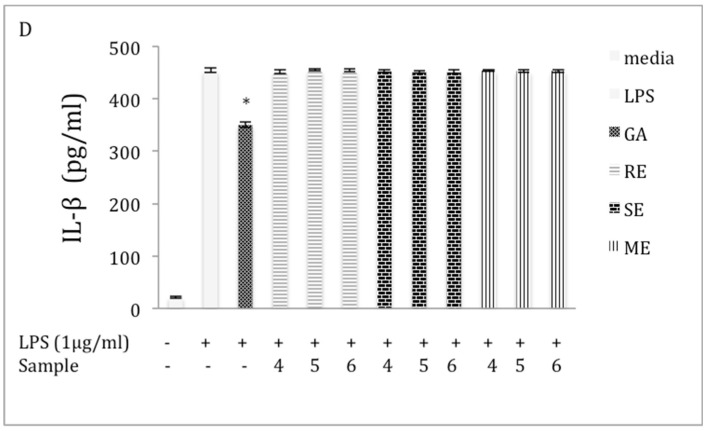
Chaga, a sclerotia formed by the Inonotus obliquus fungus, has been widely recognized for a number of medicinal properties. Although numerous scientific investigations have been published describing various biological activities of chaga from different geographical locations, little work has focused on chaga harvested in the USA or extraction techniques to maximize anti-inflammatory properties. The aim of this study was to investigate the anti-inflammatory properties of chaga collected in Maine (USA) extracted using traditional aqueous (hot water steeping) methods against lipopolysaccharide (LPS)-induced RAW 264.7 macrophages. Chaga extracts obtained from both conventional (ethanol/water) extraction methods and an accelerated solvent extraction method (ASE) at optimized conditions were compared to aqueous extracts (tea) obtained from chaga in the powder form (P) and powder form in tea bags (B) based on their effect on both nitric oxide (NO) production and pro-inflammatory cytokine expression, in particular, the expression of TNF-α, interleukin-6 (IL-6), and interleukin-β (IL-1β). Phenolic acid extracts from chaga and individual phenolic acid standards were also investigated for their effect on the same parameters. Results indicated that various chaga extracts have significant anti-inflammatory activity on LPS-stimulated RAW 264.7 cells. The inhibitory effect was through a decrease in the production of NO and the downregulation of TNF-α, IL-6, and IL-1β in RAW 264.7 macrophages. ASE1 (novel, optimized ethanol/water extraction) and P6 (six-minute steeping of powder in 100 °C water) extracts showed the highest inhibitory activity on NO production and on the expression of the inflammatory cytokines, compared to extracts obtained by conventional extraction methods.
Keywords: healthful fungal products; inflammation treatment; natural product extracts; nutraceuticals.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Taofiq O., Martins A., Barreiro M.F., Ferreira I.C. Anti-inflammatory potential of mushroom extracts and isolated metabolites. Trends Food Sci. Technol. 2016;50:193–210. doi: 10.1016/j.tifs.2016.02.005. - DOI
-
- Garlanda C., Di Liberto D., Vecchi A., La Manna M.P., Buracchi C., Caccamo N., Salerno A., Dieli F., Mantovani A. Damping excessive inflammation and tissue damage in Mycobacterium tuberculosis infection by Toll IL-1 receptor 8/single Ig IL-1-related receptor, a negative regulator of IL-1/TLR signaling. J. Immunol. 2007;179:3119–3125. doi: 10.4049/jimmunol.179.5.3119. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
