

A truncated reverse transcriptase enhances prime editing by split AAV vectors
- PMID: 35808824
- PMCID: PMC9481986
- DOI: 10.1016/j.ymthe.2022.07.001
A truncated reverse transcriptase enhances prime editing by split AAV vectors
Abstract
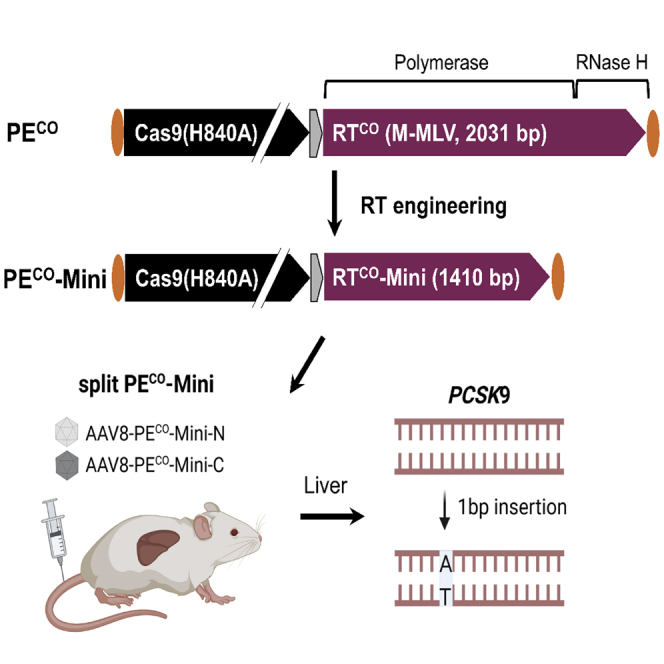
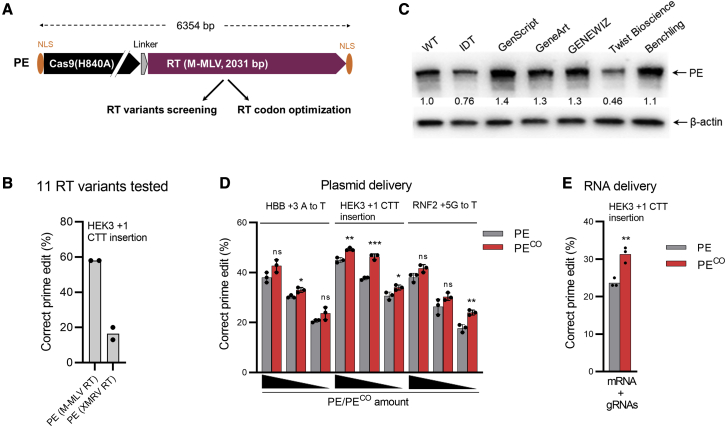
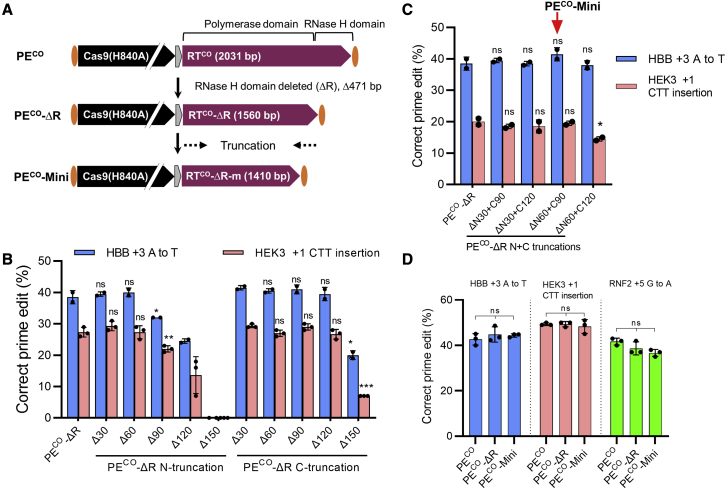
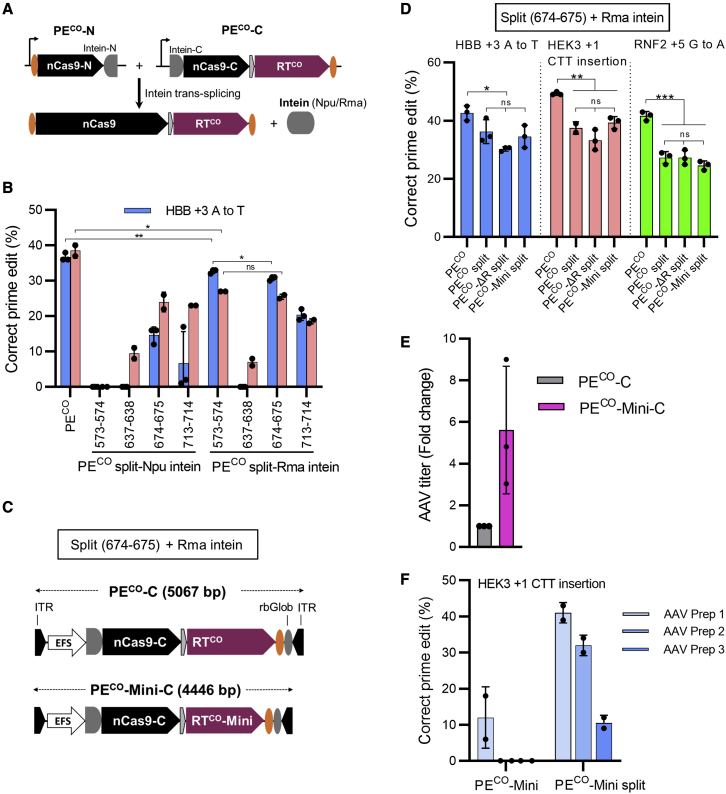
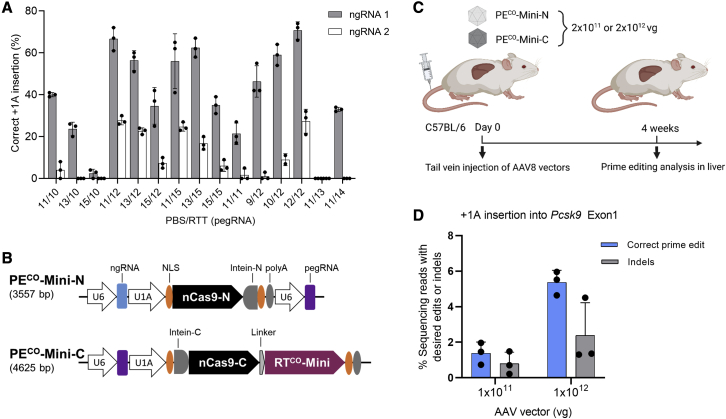
Prime editing is a new CRISPR-based, genome-editing technology that relies on the prime editor (PE), a fusion protein of Cas9-nickase and M-MLV reverse transcriptase (RT), and a prime editing guide RNA (pegRNA) that serves both to target PE to the desired genomic locus and to carry the edit to be introduced. Here, we make advancements to the RT moiety to improve prime editing efficiencies and truncations to mitigate issues with adeno-associated virus (AAV) viral vector size limitations, which currently do not support efficient delivery of the large prime editing components. These efforts include RT variant screening, codon optimization, and PE truncation by removal of the RNase H domain and further trimming. This led to a codon-optimized and size-minimized PE that has an expression advantage (1.4-fold) and size advantage (621 bp shorter). In addition, we optimize the split intein PE system and identify Rma-based Cas9 split sites (573-574 and 673-674) that combined with the truncated PE delivered by dual AAVs result in superior AAV titer and prime editing efficiency. We also show that this minimized PE gives rise to superior lentiviral vector titers (46-fold) over the regular PE in an all-in-one PE lentiviral vector. We finally deliver the minimized PE to mouse liver by dual AAV8 vectors and show up to 6% precise editing of the PCSK9 gene, thereby demonstrating the value of this truncated split PE system for in vivo applications.
Keywords: AAV vectors; CRISPR-Cas9; PASTE; gene editing; gene therapy; in vivo delivery; prime editing; reverse transcriptase.
Copyright © 2022 The American Society of Gene and Cell Therapy. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare the following competing interests: R.O.B. holds equity in Graphite Bio and UNIKUM Tx. R.O.B. is a part-time employee in UNIKUM Tx. None of the companies were involved in the present study. The remaining authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
