

DAF-2/insulin IGF-1 receptor regulates motility during aging by integrating opposite signaling from muscle and neuronal tissues
- PMID: 35808897
- PMCID: PMC9381905
- DOI: 10.1111/acel.13660
DAF-2/insulin IGF-1 receptor regulates motility during aging by integrating opposite signaling from muscle and neuronal tissues
Abstract
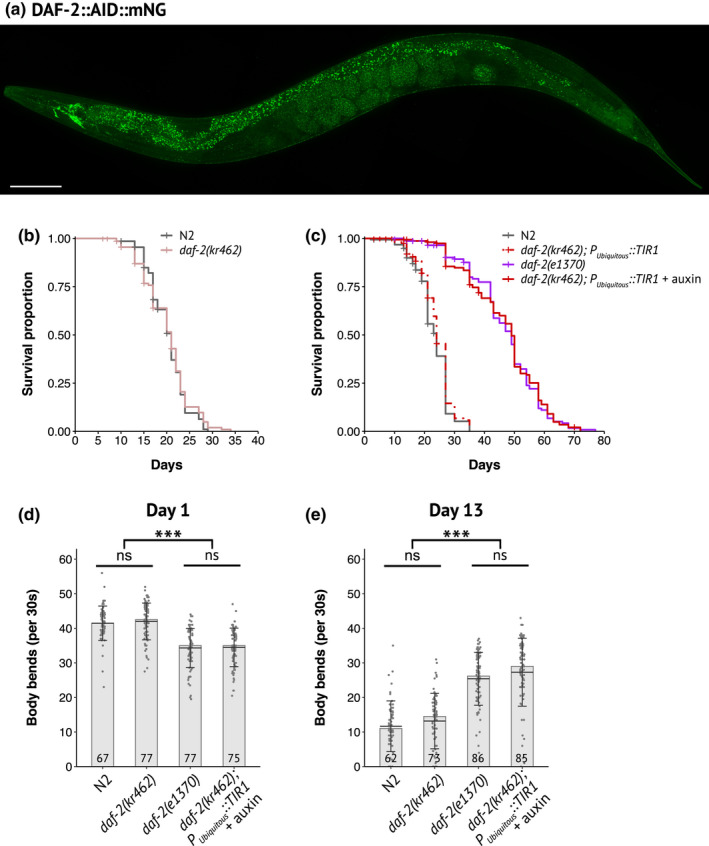
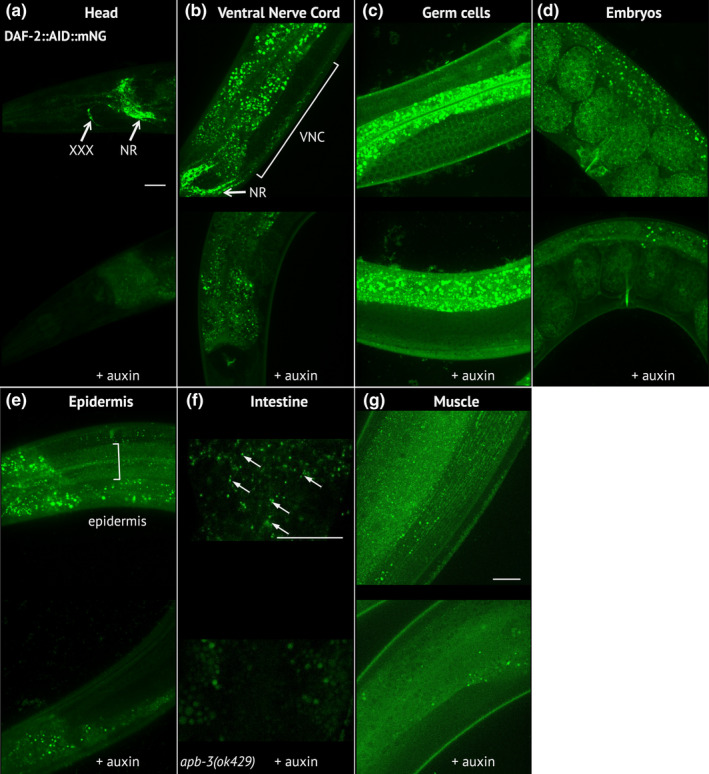
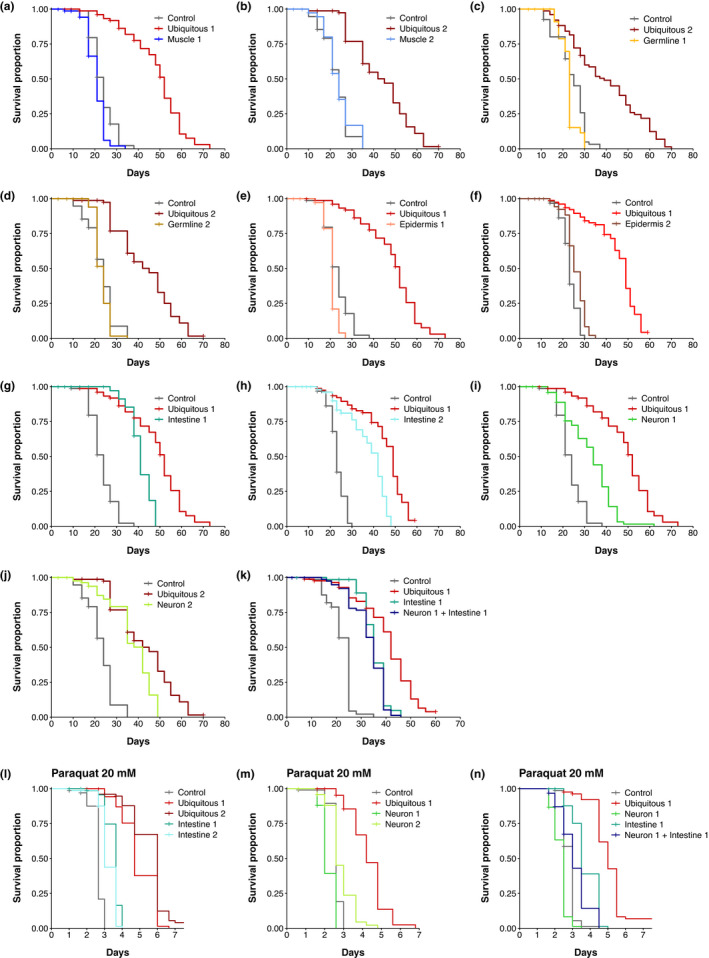
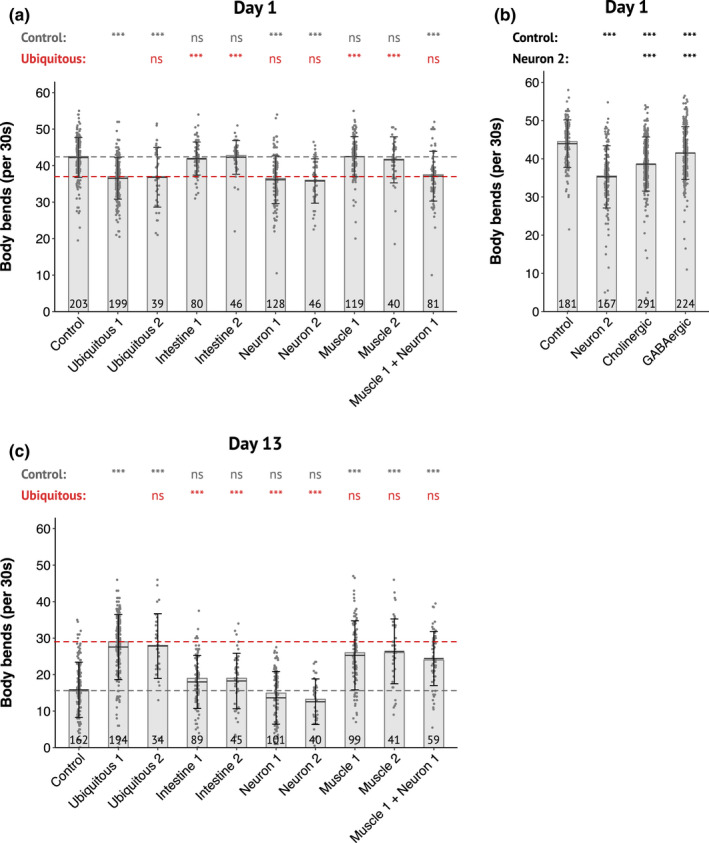
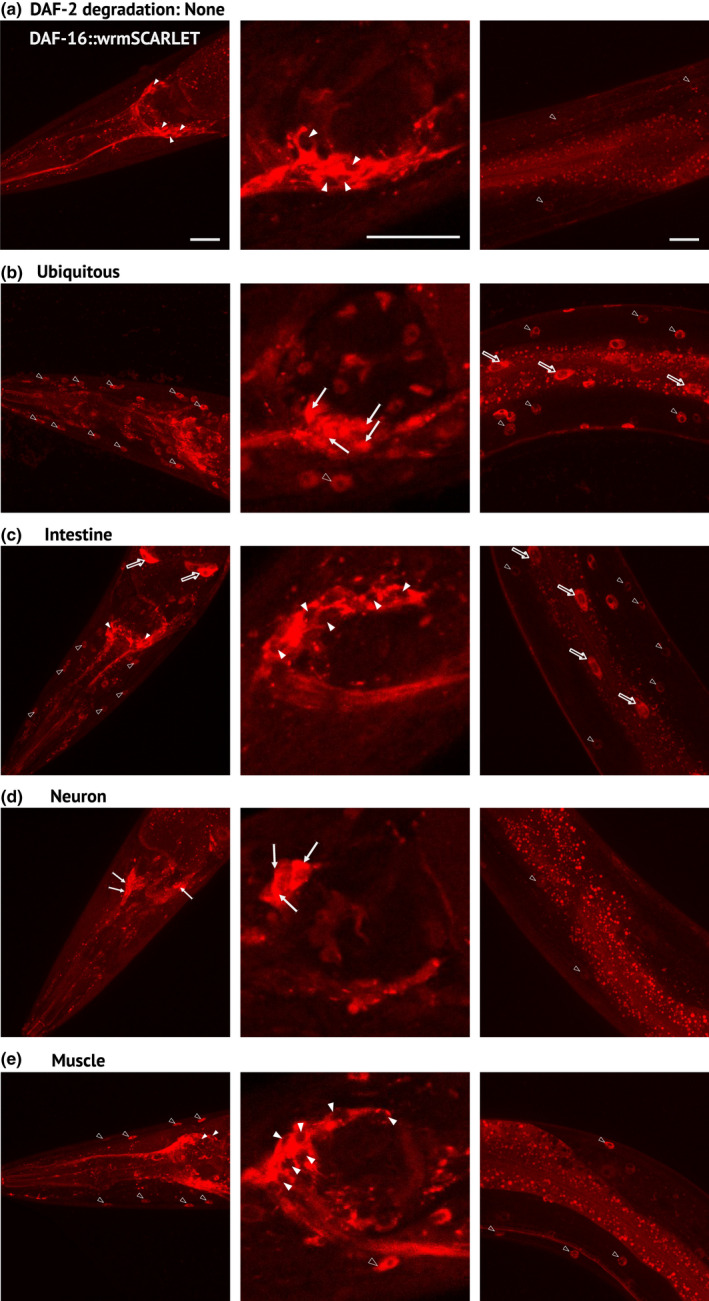
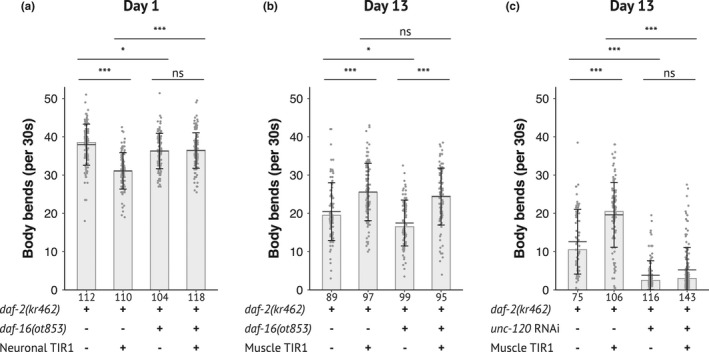
During aging, preservation of locomotion is generally considered an indicator of sustained good health, in elderlies and in animal models. In Caenorhabditis elegans, mutants of the insulin-IGF-1 receptor DAF2/IIRc represent a paradigm of healthy aging, as their increased lifespan is accompanied by a delay in age-related loss of motility. Here, we investigated the DAF-2/IIRc-dependent relationship between longevity and motility using an auxin-inducible degron to trigger tissue-specific degradation of endogenous DAF-2/IIRc. As previously reported, inactivation of DAF-2/IIRc in neurons or intestine was sufficient to extend the lifespan of worms, whereas depletion in epidermis, germline, or muscle was not. However, neither intestinal nor neuronal depletion of DAF-2/IIRc prevented the age-related loss of motility. In 1-day-old adults, DAF-2/IIRc depletion in neurons reduced motility in a DAF-16/FOXO dependent manner, while muscle depletion had no effect. By contrast, DAF-2 depletion in the muscle of middle-age animals improved their motility independently of DAF-16/FOXO but required UNC-120/SRF. Yet, neuronal or muscle DAF-2/IIRc depletion both preserved the mitochondria network in aging muscle. Overall, these results show that the motility pattern of daf-2 mutants is determined by the sequential and opposing impact of neurons and muscle tissues and can be dissociated from the regulation of the lifespan. This work also provides the characterization of a versatile tool to analyze the tissue-specific contribution of insulin-like signaling in integrated phenotypes at the whole organism level.
Keywords: daf-2; DAF-16/FOXO; UNC-120/SRF; insulin/IGF-1 signaling; lifespan; mitochondria; motility; oxidative stress.
© 2022 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Boothe, T. , Lim, G. E. , Cen, H. , Skovsø, S. , Piske, M. , Li, S. N. , Nabi, I. R. , Gilon, P. , & Johnson, J. D. (2016). Inter‐domain tagging implicates caveolin‐1 in insulin receptor trafficking and Erk signaling bias in pancreatic beta‐cells. Molecular Metabolism, 5, 366–378. 10.1016/j.molmet.2016.01.009 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
