

Functional mapping of the N-terminal region of the yeast moonlighting protein Sis2/Hal3 reveals crucial residues for Ppz1 regulation
- PMID: 35811492
- PMCID: PMC10084417
- DOI: 10.1111/febs.16572
Functional mapping of the N-terminal region of the yeast moonlighting protein Sis2/Hal3 reveals crucial residues for Ppz1 regulation
Abstract
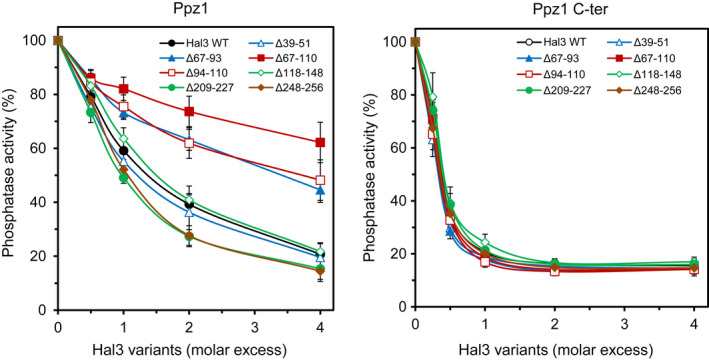
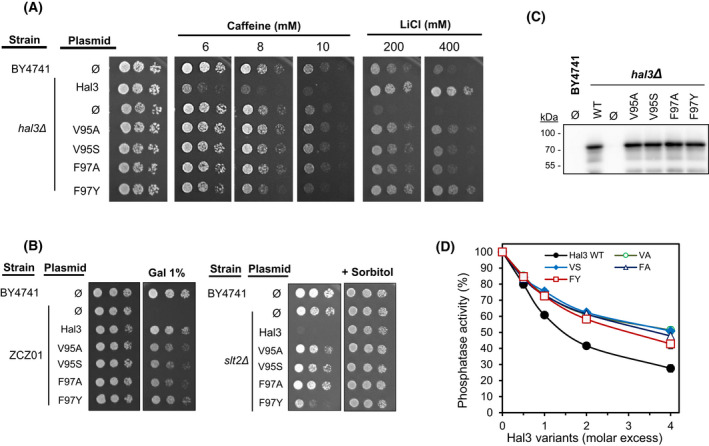
The function of the Saccharomyces cerevisiae Ppz1 phosphatase is controlled by its inhibitory subunit Hal3. Hal3 is a moonlighting protein, which associates with Cab3 to form a decarboxylase involved in the CoA biosynthetic pathway. Hal3 is composed by a conserved core PD region, required for both Ppz1 regulation and CoA biosynthesis, a long N-terminal extension, and an acidic C-terminal tail. Cab3 has a similar structure, but it is not a Ppz1 inhibitor. We show here that deletion or specific mutations in a short region of the N-terminal extension of Hal3 compromise its function as a Ppz1 inhibitor in vivo and in vitro without negatively affecting its ability to interact with the phosphatase. This study defines a R-K-X(3) -VTFS- sequence whose presence explains the unexpected ability of Cab3 (but not Hal3) to regulate Ppz1 function in Candida albicans. This sequence is conserved in a subset of fungi and it could serve to estimate the relevance of Hal3 or Cab3 proteins in regulating fungal Ppz enzymes. We also show that the removal of the motif moderately affects both Ppz1 intracellular relocalization and counteraction of toxicity in cells overexpressing the phosphatase. Thus, our work contributes to our understanding of the regulation of Ppz phosphatases, which are determinants for virulence in some pathogenic fungi.
Keywords: disordered regions; mutagenesis; protein phosphatase; regulatory subunit; yeasts.
© 2022 The Authors. The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








Similar articles
-
Fungal Hal3 (and Its Close Relative Cab3) as Moonlighting Proteins.J Fungi (Basel). 2022 Oct 11;8(10):1066. doi: 10.3390/jof8101066. J Fungi (Basel). 2022. PMID: 36294631 Free PMC article. Review.
-
Functional characterization of the yeast Ppz1 phosphatase inhibitory subunit Hal3: a mutagenesis study.J Biol Chem. 2004 Oct 8;279(41):42619-27. doi: 10.1074/jbc.M405656200. Epub 2004 Jul 29. J Biol Chem. 2004. PMID: 15292171
-
Mutations at the hydrophobic core affect Hal3 trimer stability, reducing its Ppz1 inhibitory capacity but not its PPCDC moonlighting function.Sci Rep. 2018 Oct 2;8(1):14701. doi: 10.1038/s41598-018-32979-x. Sci Rep. 2018. PMID: 30279472 Free PMC article.
-
The toxic effects of yeast Ppz1 phosphatase are counteracted by subcellular relocalization mediated by its regulatory subunit Hal3.FEBS Lett. 2022 Jun;596(12):1556-1566. doi: 10.1002/1873-3468.14330. Epub 2022 Mar 23. FEBS Lett. 2022. PMID: 35278214
-
When Phosphatases Go Mad: The Molecular Basis for Toxicity of Yeast Ppz1.Int J Mol Sci. 2022 Apr 13;23(8):4304. doi: 10.3390/ijms23084304. Int J Mol Sci. 2022. PMID: 35457140 Free PMC article. Review.
Cited by
-
Fungal Hal3 (and Its Close Relative Cab3) as Moonlighting Proteins.J Fungi (Basel). 2022 Oct 11;8(10):1066. doi: 10.3390/jof8101066. J Fungi (Basel). 2022. PMID: 36294631 Free PMC article. Review.
References
-
- Brautigan DL. Protein ser/Thr phosphatases–the ugly ducklings of cell signalling. FEBS J. 2013;280:324–5. - PubMed
-
- Posas F, Casamayor A, Morral N, Ariño J. Molecular cloning and analysis of a yeast protein phosphatase with an unusual amino‐terminal region. J Biol Chem. 1992;267:11734–40. - PubMed
-
- Posas F, Camps M, Ariño J. The PPZ protein phosphatases are important determinants of salt tolerance in yeast cells. J Biol Chem. 1995;270:13036–41. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
