

Birdsong and the Neural Regulation of Positive Emotion
- PMID: 35814050
- PMCID: PMC9258629
- DOI: 10.3389/fpsyg.2022.903857
Birdsong and the Neural Regulation of Positive Emotion
Abstract
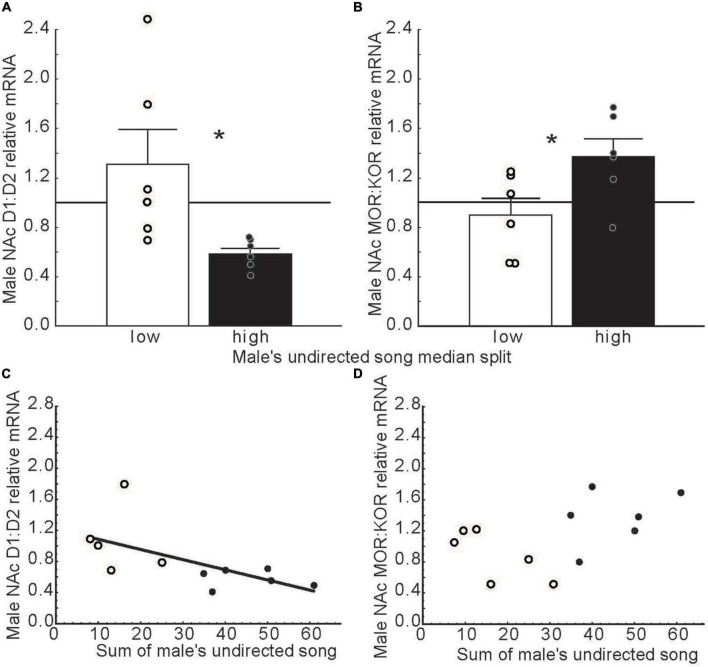
Birds are not commonly admired for emotional expression, and when they are, the focus is typically on negative states; yet vocal behavior is considered a direct reflection of an individual's emotional state. Given that over 4000 species of songbird produce learned, complex, context-specific vocalizations, we make the case that songbirds are conspicuously broadcasting distinct positive emotional states and that hearing songs can also induce positive states in other birds. Studies are reviewed that demonstrate that that the production of sexually motivated song reflects an emotional state of anticipatory reward-seeking (i.e., mate-seeking), while outside the mating context song in gregarious flocks reflects a state of intrinsic reward. Studies are also reviewed that demonstrate that hearing song induces states of positive anticipation and reward. This review brings together numerous studies that highlight a potentially important role for the songbird nucleus accumbens, a region nearly synonymous with reward in mammals, in positive emotional states that underlie singing behavior and responses to song. It is proposed that the nucleus accumbens is part of an evolutionarily conserved circuitry that contributes context-dependently to positive emotional states that motivate and reward singing behavior and responses to song. Neural mechanisms that underlie basic emotions appear to be conserved and similar across vertebrates. Thus, these findings in songbirds have the potential to provide insights into interventions that can restore positive social interactions disrupted by mental health disorders in humans.
Keywords: communication; emotion; motivation; nucleus accumbens (NAc); positive affect; reward; songbird; vocal behavior.
Copyright © 2022 Riters, Polzin, Maksimoski, Stevenson and Alger.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures








References
-
- Adret P. (1993). Operant conditioning, song learning and imprinting to taped song in the zebra finch. Anim. Behav. 46 149–159.
-
- Akins C. K. (1998). Context excitation and modulation of conditioned sexual behavior. Anim. Learn. Behav. 26 416–426.
-
- Alger S. J., Riters L. V. (2006). Lesions to the medial preoptic nucleus differentially affect singing and nest box-directed behaviors within and outside of the breeding season in European starlings (Sturnus vulgaris). Behav. Neurosci. 120 1326–1336. 10.1037/0735-7044.120.6.1326 - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources