

Local BMP signaling: A sensor for synaptic activity that balances synapse growth and function
- PMID: 35817503
- PMCID: PMC11102767
- DOI: 10.1016/bs.ctdb.2022.04.001
Local BMP signaling: A sensor for synaptic activity that balances synapse growth and function
Abstract
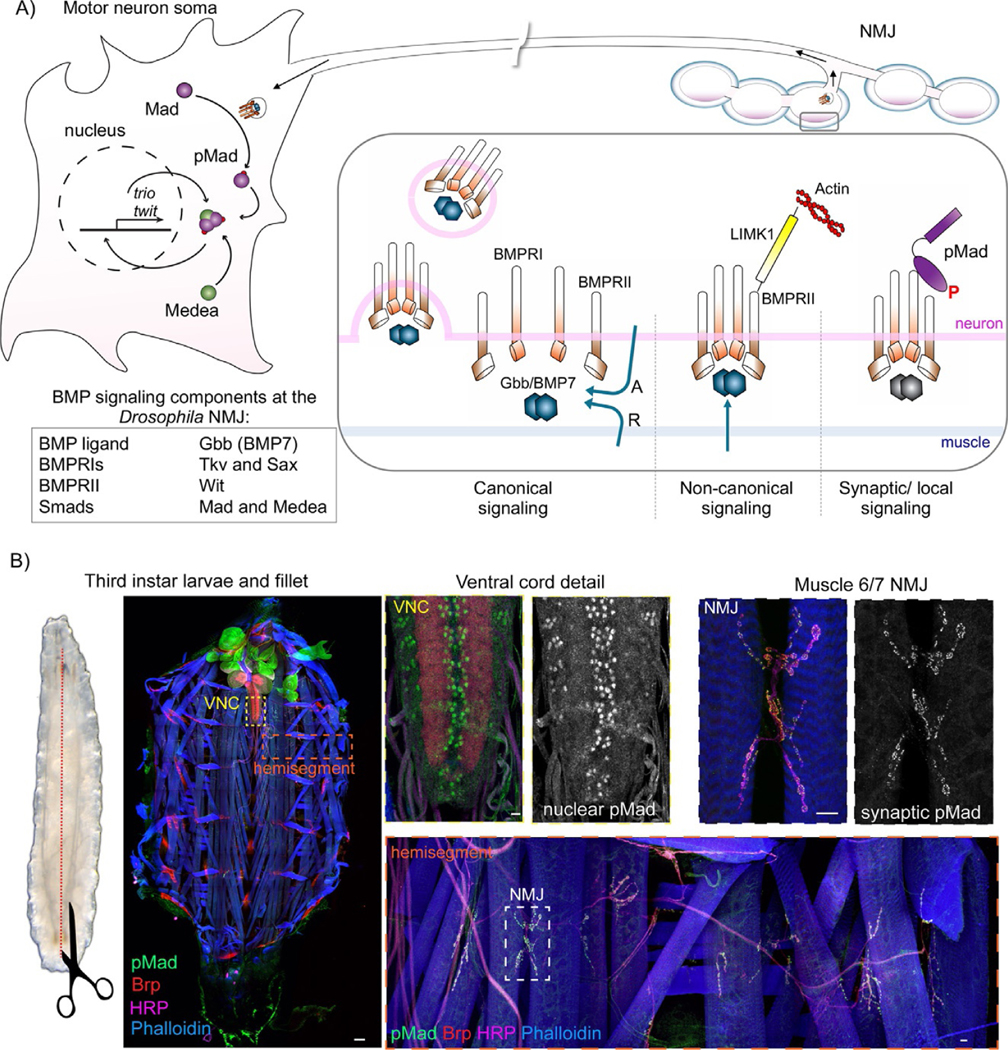
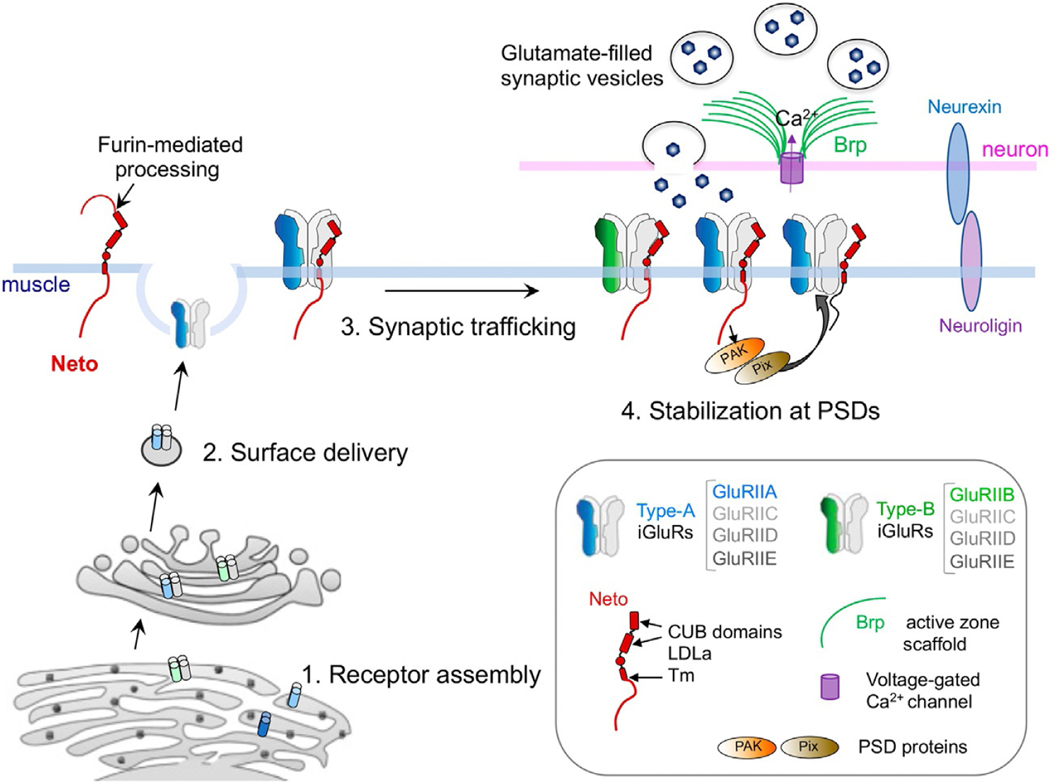
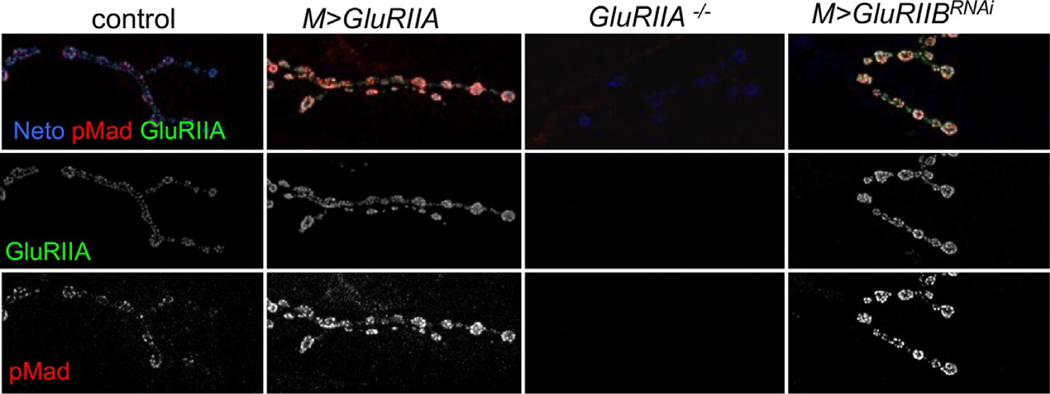
Synapse development is coordinated by intercellular communication between the pre- and postsynaptic compartments, and by neuronal activity itself. In flies as in vertebrates, neuronal activity induces input-specific changes in the synaptic strength so that the entire circuit maintains stable function in the face of many challenges, including changes in synapse number and strength. But how do neurons sense synapse activity? In several studies carried out using the Drosophila neuromuscular junction (NMJ), we demonstrated that local BMP signaling provides an exquisite sensor for synapse activity. Here we review the main features of this exquisite sensor and discuss its functioning beyond monitoring the synapse activity but rather as a key controller that operates in coordination with other BMP signaling pathways to balance synapse growth, maturation and function.
Keywords: BMP signaling; Local BMP signaling; Positive feedback loop; Sensor for synapse activity; Synapse assembly.
Copyright © 2022 Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Postsynaptic glutamate receptors regulate local BMP signaling at the Drosophila neuromuscular junction.Development. 2014 Jan;141(2):436-47. doi: 10.1242/dev.097758. Epub 2013 Dec 18. Development. 2014. PMID: 24353060 Free PMC article.
-
Retrograde BMP signaling at the synapse: a permissive signal for synapse maturation and activity-dependent plasticity.J Neurosci. 2013 Nov 6;33(45):17937-50. doi: 10.1523/JNEUROSCI.6075-11.2013. J Neurosci. 2013. PMID: 24198381 Free PMC article.
-
Neuroligin 4 regulates synaptic growth via the bone morphogenetic protein (BMP) signaling pathway at the Drosophila neuromuscular junction.J Biol Chem. 2017 Nov 3;292(44):17991-18005. doi: 10.1074/jbc.M117.810242. Epub 2017 Sep 14. J Biol Chem. 2017. PMID: 28912273 Free PMC article.
-
Synapse development and maturation at the drosophila neuromuscular junction.Neural Dev. 2020 Aug 2;15(1):11. doi: 10.1186/s13064-020-00147-5. Neural Dev. 2020. PMID: 32741370 Free PMC article. Review.
-
Synaptic homeostats: latent plasticity revealed at the Drosophila neuromuscular junction.Cell Mol Life Sci. 2021 Apr;78(7):3159-3179. doi: 10.1007/s00018-020-03732-3. Epub 2021 Jan 15. Cell Mol Life Sci. 2021. PMID: 33449150 Free PMC article. Review.
Cited by
-
Neto proteins differentially modulate the gating properties of Drosophila NMJ glutamate receptors.bioRxiv [Preprint]. 2024 Apr 26:2024.04.22.590603. doi: 10.1101/2024.04.22.590603. bioRxiv. 2024. Update in: J Physiol. 2024 Dec;602(24):7043-7064. doi: 10.1113/JP287331. PMID: 38903091 Free PMC article. Updated. Preprint.
-
scRNA-seq data from the larval Drosophila ventral cord provides a resource for studying motor systems function and development.Dev Cell. 2024 May 6;59(9):1210-1230.e9. doi: 10.1016/j.devcel.2024.03.016. Epub 2024 Apr 2. Dev Cell. 2024. PMID: 38569548 Free PMC article.
-
Bone morphogenetic protein signaling: the pathway and its regulation.Genetics. 2024 Feb 7;226(2):iyad200. doi: 10.1093/genetics/iyad200. Genetics. 2024. PMID: 38124338 Free PMC article. Review.
-
Mitochondrial ROS and HIF-1α signaling mediate synaptic plasticity in the critical period.PLoS Biol. 2025 Aug 13;23(8):e3003338. doi: 10.1371/journal.pbio.3003338. eCollection 2025 Aug. PLoS Biol. 2025. PMID: 40802851 Free PMC article.
-
Control of neuronal excitation-inhibition balance by BMP-SMAD1 signalling.Nature. 2024 May;629(8011):402-409. doi: 10.1038/s41586-024-07317-z. Epub 2024 Apr 17. Nature. 2024. PMID: 38632412 Free PMC article.
References
-
- Aberle H, Haghighi AP, Fetter RD, McCabe BD, Magalhaes TR, & Goodman CS (2002). wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila. Neuron, 33, 545–558. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
