

Evolutionary History of Sexual Differentiation Mechanism in Insects
- PMID: 35820410
- PMCID: PMC9290531
- DOI: 10.1093/molbev/msac145
Evolutionary History of Sexual Differentiation Mechanism in Insects
Abstract
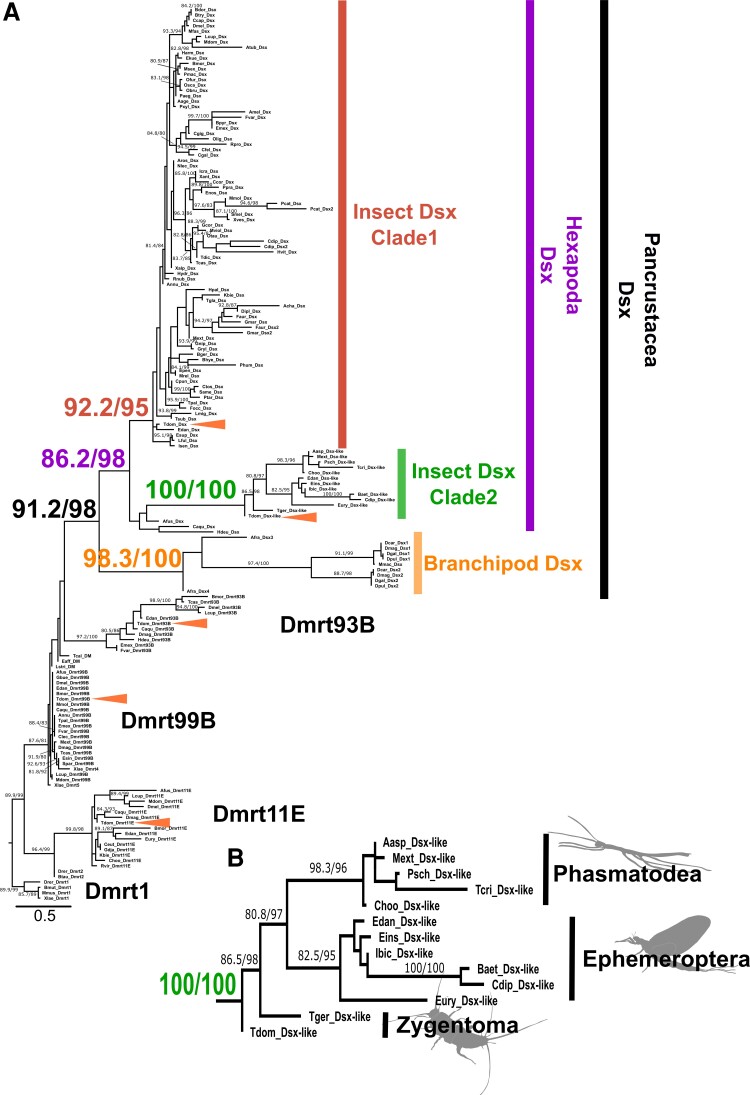
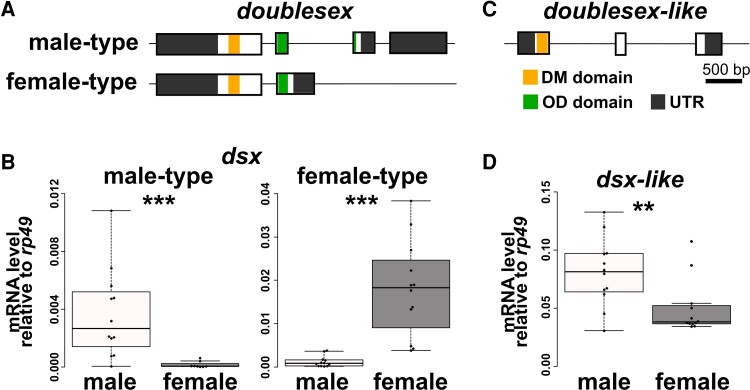
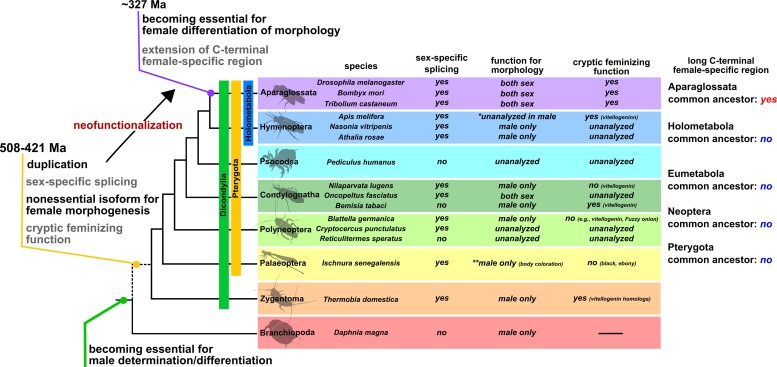
Alternative splicing underpins functional diversity in proteins and the complexity and diversity of eukaryotes. An example is the doublesex gene, the key transcriptional factor in arthropod sexual differentiation. doublesex is controlled by sex-specific splicing and promotes both male and female differentiation in holometabolan insects, whereas in hemimetabolan species, doublesex has sex-specific isoforms but is not required for female differentiation. How doublesex evolved to be essential for female development remains largely unknown. Here, we investigate ancestral states of doublesex using Thermobia domestica belonging to Zygentoma, the sister group of Pterygota, that is, winged insects. We find that, in T. domestica, doublesex expresses sex-specific isoforms but is only necessary for male differentiation of sexual morphology. This result supports the hypothesis that doublesex initially promoted male differentiation during insect evolution. However, T. domestica doublesex has a short female-specific region and upregulates the expression of vitellogenin homologs in females, suggesting that doublesex may already play some role in female morphogenesis of the common ancestor of Pterygota. Reconstruction of the ancestral sequence and prediction of protein structures show that the female-specific isoform of doublesex has an extended C-terminal disordered region in holometabolan insects but not in nonholometabolan species. We propose that doublesex acquired its function in female morphogenesis through a change in the protein motif structure rather than the emergence of the female-specific exon.
Keywords: doublesex; Zygentoma; alternative splicing; insect; sexual differentiation.
© The Author(s) 2022. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures




Similar articles
-
Transgenic and knockout analyses of Masculinizer and doublesex illuminated the unique functions of doublesex in germ cell sexual development of the silkworm, Bombyx mori.BMC Dev Biol. 2020 Sep 21;20(1):19. doi: 10.1186/s12861-020-00224-2. BMC Dev Biol. 2020. PMID: 32957956 Free PMC article.
-
Characterization of the doublesex gene within the Culex pipiens complex suggests regulatory plasticity at the base of the mosquito sex determination cascade.BMC Evol Biol. 2015 Jun 11;15:108. doi: 10.1186/s12862-015-0386-1. BMC Evol Biol. 2015. PMID: 26058583 Free PMC article.
-
Genomic organization and splicing evolution of the doublesex gene, a Drosophila regulator of sexual differentiation, in the dengue and yellow fever mosquito Aedes aegypti.BMC Evol Biol. 2011 Feb 10;11:41. doi: 10.1186/1471-2148-11-41. BMC Evol Biol. 2011. PMID: 21310052 Free PMC article.
-
Doublesex: a conserved downstream gene controlled by diverse upstream regulators.J Genet. 2010 Sep;89(3):341-56. doi: 10.1007/s12041-010-0046-6. J Genet. 2010. PMID: 20877001 Review.
-
Structure, function and evolution of sex-determining systems in Dipteran insects.Development. 2000 Feb;127(4):667-77. doi: 10.1242/dev.127.4.667. Development. 2000. PMID: 10648226 Review.
Cited by
-
Recognition of polymorphic Csd proteins determines sex in the honeybee.Sci Adv. 2023 Oct 6;9(40):eadg4239. doi: 10.1126/sciadv.adg4239. Epub 2023 Oct 4. Sci Adv. 2023. PMID: 37792946 Free PMC article.
-
Developmental transcriptomics of the Firebrat: Exploring developmental expression patterns and morphology during the embryogenesis of Thermobia domestica.PLoS One. 2025 Jun 5;20(6):e0324844. doi: 10.1371/journal.pone.0324844. eCollection 2025. PLoS One. 2025. PMID: 40471973 Free PMC article.
-
The function and evolution of a genetic switch controlling sexually dimorphic eye differentiation in honeybees.Nat Commun. 2023 Jan 28;14(1):463. doi: 10.1038/s41467-023-36153-4. Nat Commun. 2023. PMID: 36709321 Free PMC article.
-
Analyses of the Dmrt family in a decapod crab, Eriocheir sinensis uncover new facets on the evolution of DM domain genes.Front Physiol. 2023 May 26;14:1201846. doi: 10.3389/fphys.2023.1201846. eCollection 2023. Front Physiol. 2023. PMID: 37304820 Free PMC article.
-
Dedicated developmental programing for group-supporting behaviors in eusocial honeybees.Sci Adv. 2024 Nov;10(44):eadp3953. doi: 10.1126/sciadv.adp3953. Epub 2024 Nov 1. Sci Adv. 2024. PMID: 39485851 Free PMC article.
References
-
- R Core Team. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing.
-
- An W, Wensink PC. 1995. Integrating sex-and tissue-specific regulation within a single Drosophila enhancer. Genes Dev. 9(2):256–266. - PubMed
-
- Bear A, Monteiro A. 2013. Both cell-autonomous mechanisms and hormones contribute to sexual development in vertebrates and insects. Bioessays 35(8):725–732. - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
