

A new paradigm for regulation of protein phosphatase 2A function via Src and Fyn kinase-mediated tyrosine phosphorylation
- PMID: 35820485
- PMCID: PMC9396060
- DOI: 10.1016/j.jbc.2022.102248
A new paradigm for regulation of protein phosphatase 2A function via Src and Fyn kinase-mediated tyrosine phosphorylation
Abstract
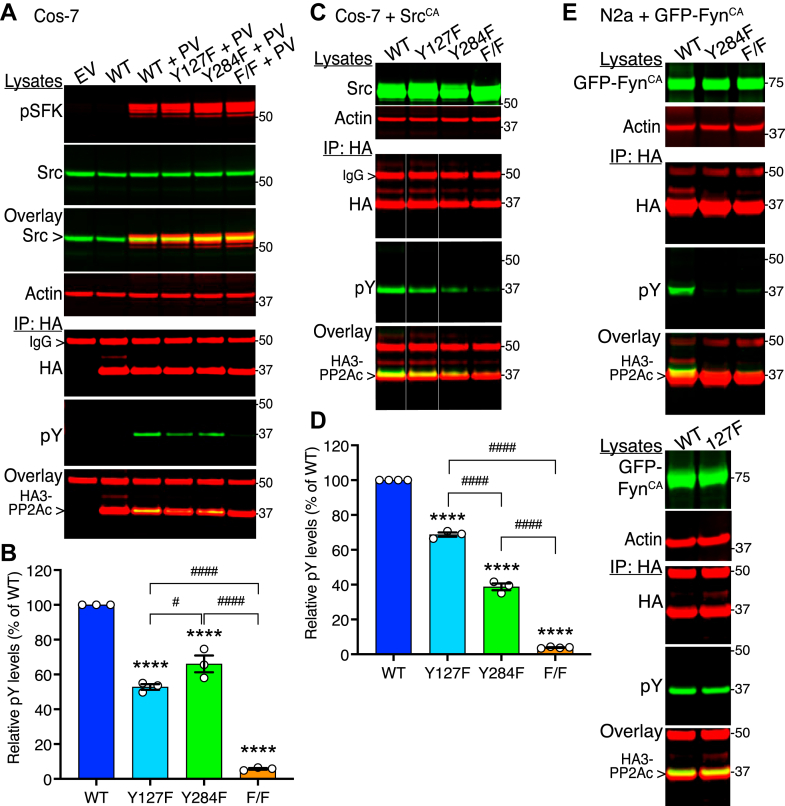
Protein phosphatase 2A (PP2A) is a major phospho-Ser/Thr phosphatase and a key regulator of cellular signal transduction pathways. While PP2A dysfunction has been linked to human cancer and neurodegenerative disorders such as Alzheimer's disease (AD), PP2A regulation remains relatively poorly understood. It has been reported that the PP2A catalytic subunit (PP2Ac) is inactivated by a single phosphorylation at the Tyr307 residue by tyrosine kinases such as v-Src. However, multiple mass spectrometry studies have revealed the existence of other putative PP2Ac phosphorylation sites in response to activation of Src and Fyn, two major Src family kinases (SFKs). Here, using PP2Ac phosphomutants and novel phosphosite-specific PP2Ac antibodies, we show that cellular pools of PP2Ac are instead phosphorylated on both Tyr127 and Tyr284 upon Src activation, and on Tyr284 following Fyn activation. We found these phosphorylation events enhanced the interaction of PP2Ac with SFKs. In addition, we reveal SFK-mediated phosphorylation of PP2Ac at Y284 promotes dissociation of the regulatory Bα subunit, altering PP2A substrate specificity; the phosphodeficient Y127/284F and Y284F PP2Ac mutants prevented SFK-mediated phosphorylation of Tau at the CP13 (pSer202) epitope, a pathological hallmark of AD, and SFK-dependent activation of ERK, a major growth regulatory kinase upregulated in many cancers. Our findings demonstrate a novel PP2A regulatory mechanism that challenges the existing dogma on the inhibition of PP2A catalytic activity by Tyr307 phosphorylation. We propose dysregulation of SFK signaling in cancer and AD can lead to alterations in PP2A phosphorylation and subsequent deregulation of key PP2A substrates, including ERK and Tau.
Keywords: ERK; Fyn; PP2A; Src; Tau; protein phosphatase 2A; tyrosine phosphorylation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest B. E. W. and B. W. S. are cofounders of Turkey Creek Biotechnology LLC and have equity ownership. The authors declare no other competing conflicts of interest with the contents of this article.
Figures






References
-
- Parsons S.J., Parsons J.T. Src family kinases, key regulators of signal transduction. Oncogene. 2004;23:7906–7909. - PubMed
-
- Elias D., Ditzel H.J. Fyn is an important molecule in cancer pathogenesis and drug resistance. Pharmacol. Res. 2015;100:250–254. - PubMed
-
- Roskoski R., Jr. Src protein-tyrosine kinase structure, mechanism, and small molecule inhibitors. Pharmacol. Res. 2015;94:9–25. - PubMed
-
- Lee G. Tau and src family tyrosine kinases. Biochim. Biophys. Acta. 2005;1739:323–330. - PubMed
-
- Ittner A., Ittner L.M. Dendritic tau in Alzheimer's disease. Neuron. 2018;99:13–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
