

KARRIKIN UPREGULATED F-BOX 1 negatively regulates drought tolerance in Arabidopsis
- PMID: 35822606
- PMCID: PMC9706471
- DOI: 10.1093/plphys/kiac336
KARRIKIN UPREGULATED F-BOX 1 negatively regulates drought tolerance in Arabidopsis
Abstract
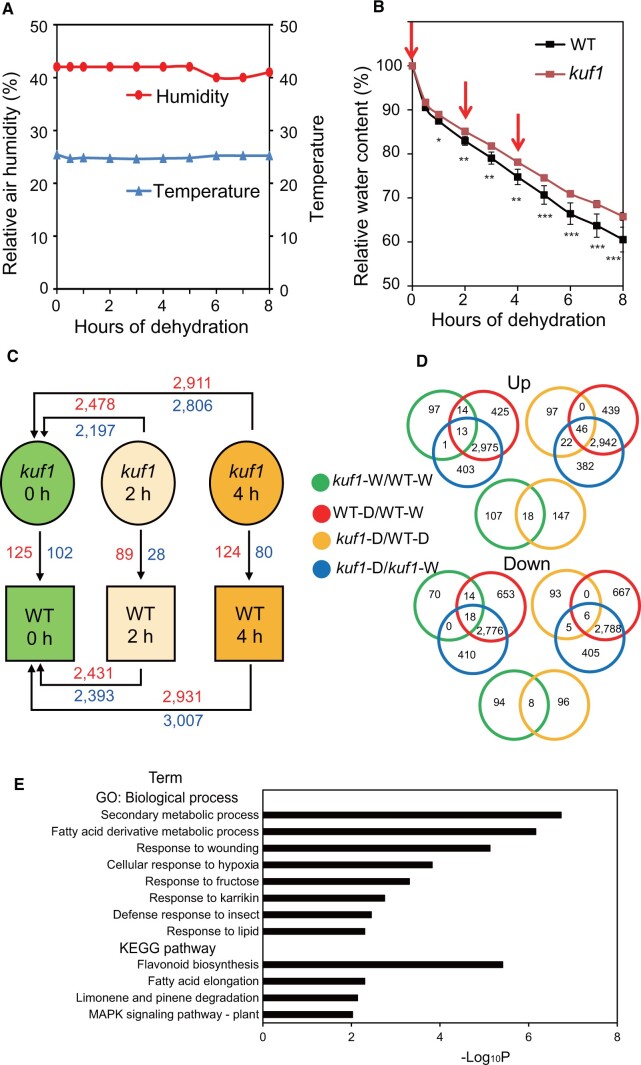
The karrikin (KAR) receptor and several related signaling components have been identified by forward genetic screening, but only a few studies have reported on upstream and downstream KAR signaling components and their roles in drought tolerance. Here, we characterized the functions of KAR UPREGULATED F-BOX 1 (KUF1) in drought tolerance using a reverse genetics approach in Arabidopsis (Arabidopsis thaliana). We observed that kuf1 mutant plants were more tolerant to drought stress than wild-type (WT) plants. To clarify the mechanisms by which KUF1 negatively regulates drought tolerance, we performed physiological, transcriptome, and morphological analyses. We found that kuf1 plants limited leaf water loss by reducing stomatal aperture and cuticular permeability. In addition, kuf1 plants showed increased sensitivity of stomatal closure, seed germination, primary root growth, and leaf senescence to abscisic acid (ABA). Genome-wide transcriptome comparisons of kuf1 and WT rosette leaves before and after dehydration showed that the differences in various drought tolerance-related traits were accompanied by differences in the expression of genes associated with stomatal closure (e.g. OPEN STOMATA 1), lipid and fatty acid metabolism (e.g. WAX ESTER SYNTHASE), and ABA responsiveness (e.g. ABA-RESPONSIVE ELEMENT 3). The kuf1 mutant plants had higher root/shoot ratios and root hair densities than WT plants, suggesting that they could absorb more water than WT plants. Together, these results demonstrate that KUF1 negatively regulates drought tolerance by modulating various physiological traits, morphological adjustments, and ABA responses and that the genetic manipulation of KUF1 in crops is a potential means of enhancing their drought tolerance.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures






References
-
- Abdelrahman M, Jogaiah S, Burritt DJ, Tran LP (2018) Legume genetic resources and transcriptome dynamics under abiotic stress conditions. Plant Cell Environ 41: 1972–1983 - PubMed
-
- Acharya BR, Jeon BW, Zhang W, Assmann SM (2013) Open Stomata 1 (OST1) is limiting in abscisic acid responses of Arabidopsis guard cells. New Phytol 200: 1049–1063 - PubMed
-
- Buckley TN (2019) How do stomata respond to water status? New Phytol 224: 21–36 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
