

SlJAZ10 and SlJAZ11 mediate dark-induced leaf senescence and regeneration
- PMID: 35830385
- PMCID: PMC9278786
- DOI: 10.1371/journal.pgen.1010285
SlJAZ10 and SlJAZ11 mediate dark-induced leaf senescence and regeneration
Abstract
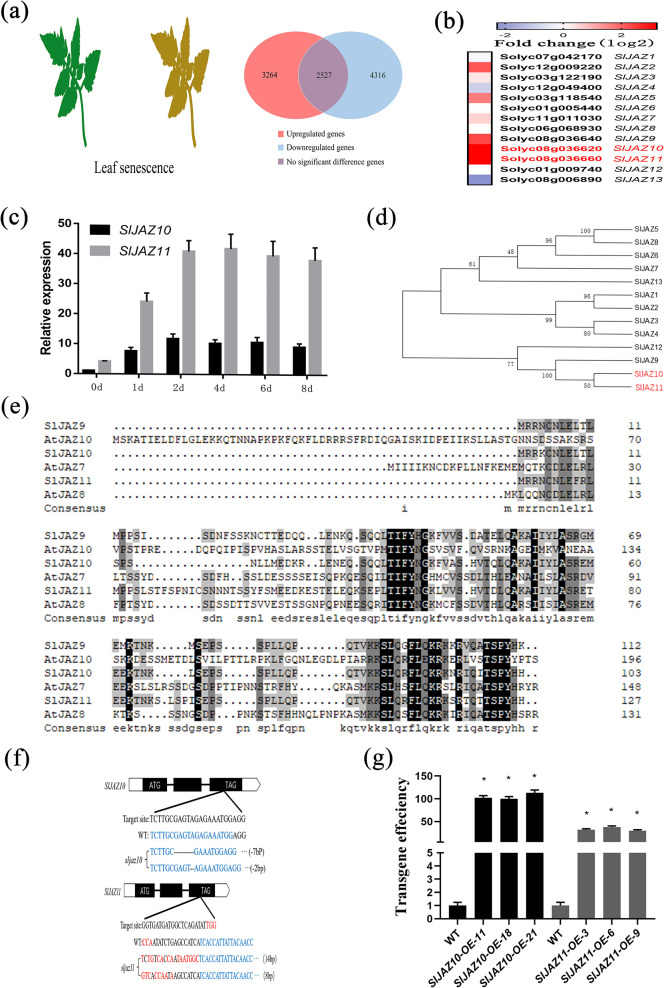
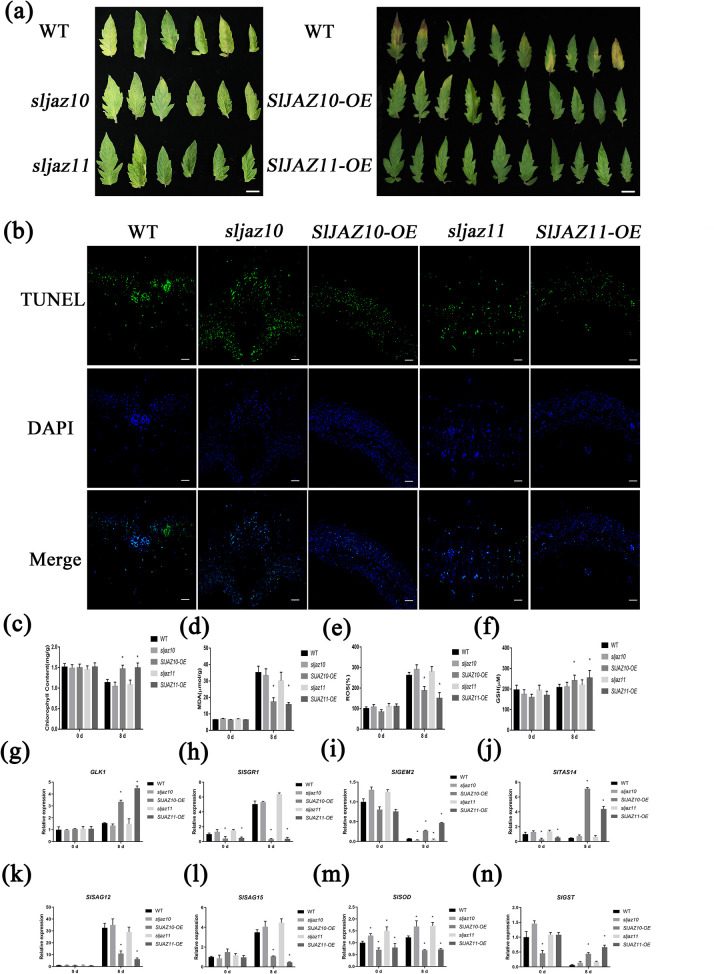
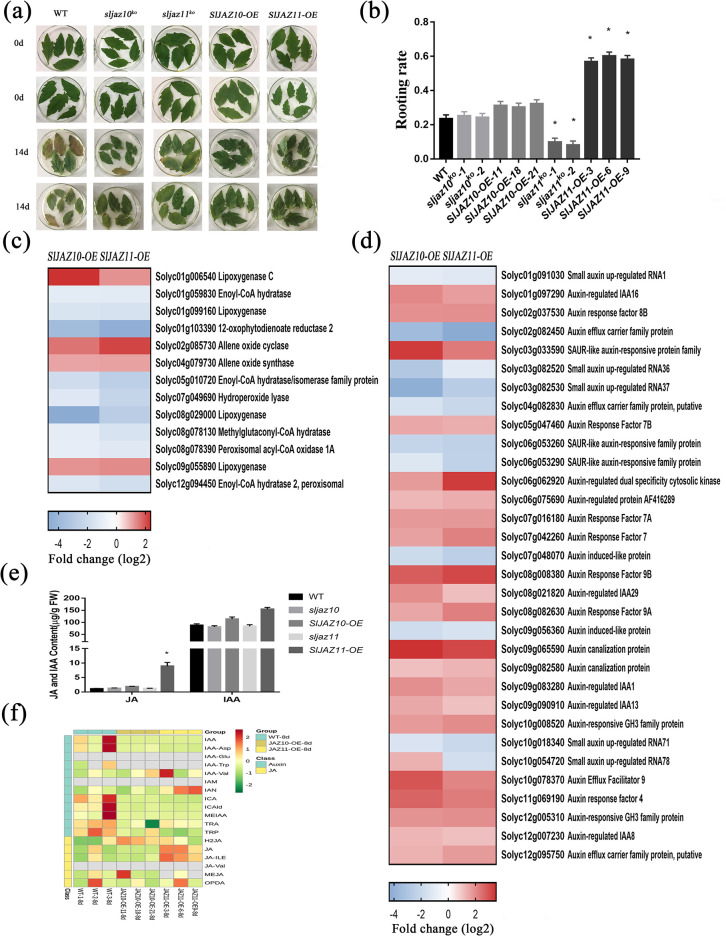
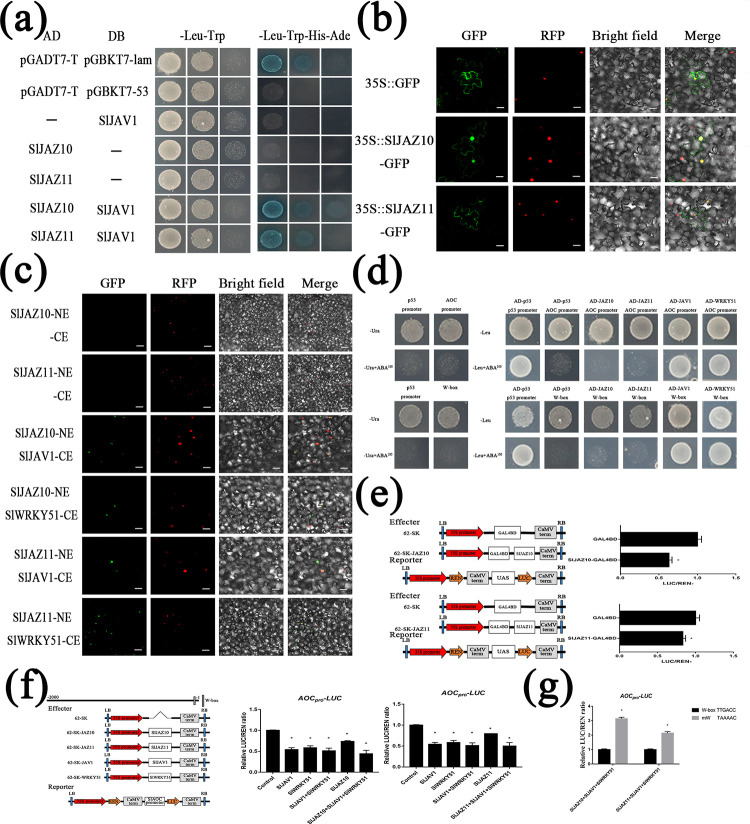
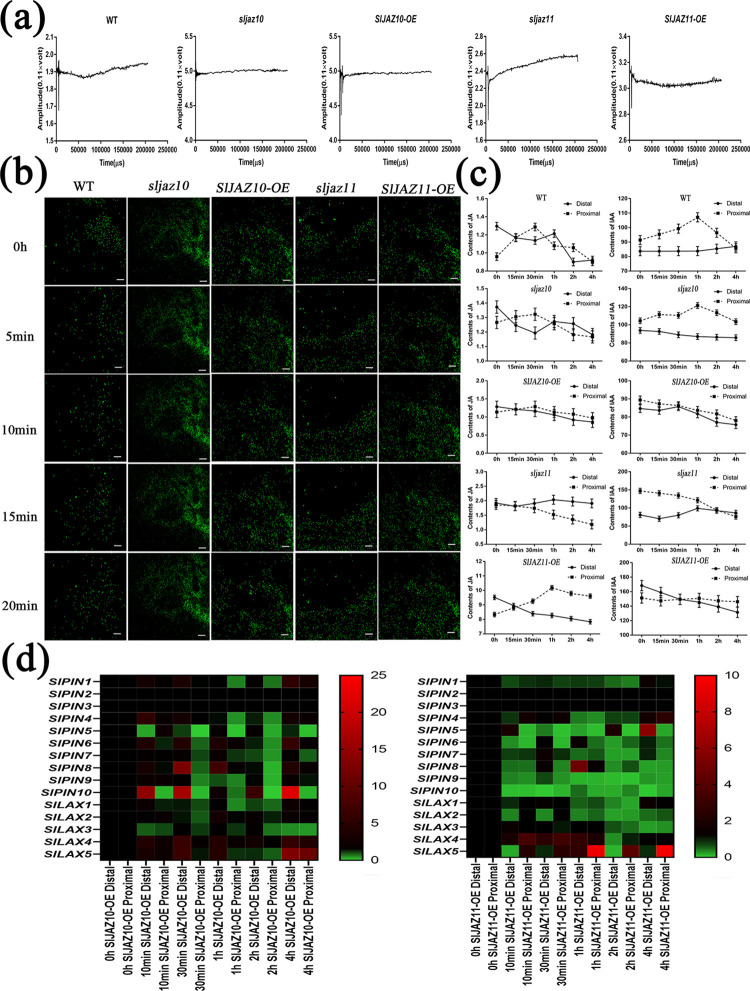
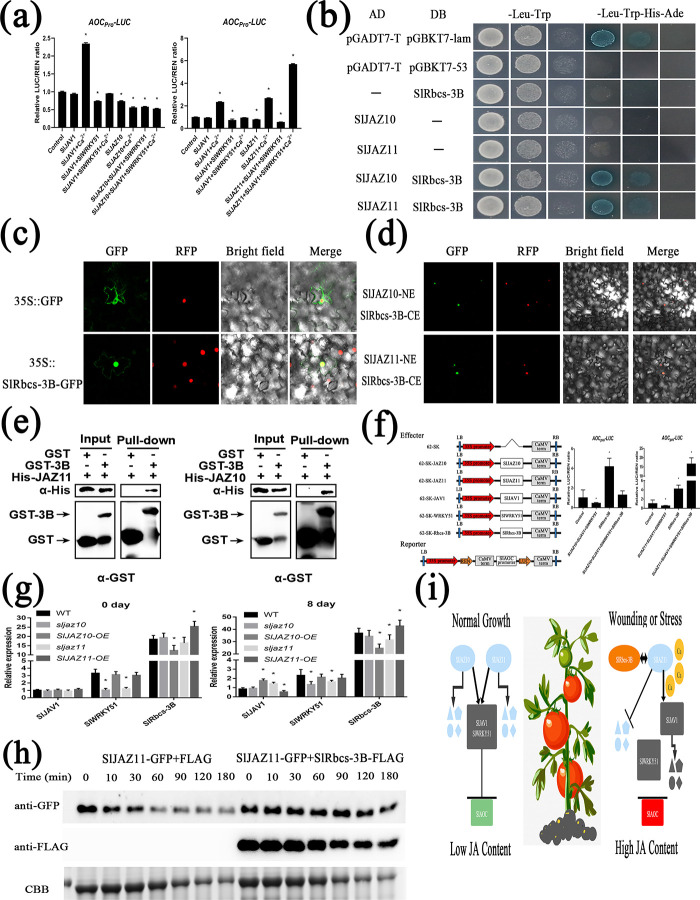
During evolutionary adaptation, the mechanisms for self-regulation are established between the normal growth and development of plants and environmental stress. The phytohormone jasmonate (JA) is a key tie of plant defence and development, and JASMONATE-ZIM DOMAIN (JAZ) repressor proteins are key components in JA signalling pathways. Here, we show that JAZ expression was affected by leaf senescence from the transcriptomic data. Further investigation revealed that SlJAZ10 and SlJAZ11 positively regulate leaf senescence and that SlJAZ11 can also promote plant regeneration. Moreover, we reveal that the SlJAV1-SlWRKY51 (JW) complex could suppress JA biosynthesis under normal growth conditions. Immediately after injury, SlJAZ10 and SlJAZ11 can regulate the activity of the JW complex through the effects of electrical signals and Ca2+ waves, which in turn affect JA biosynthesis, causing a difference in the regeneration phenotype between SlJAZ10-OE and SlJAZ11-OE transgenic plants. In addition, SlRbcs-3B could maintain the protein stability of SlJAZ11 to protect it from degradation. Together, SlJAZ10 and SlJAZ11 not only act as repressors of JA signalling to leaf senescence, but also regulate plant regeneration through coordinated electrical signals, Ca2+ waves, hormones and transcriptional regulation. Our study provides critical insights into the mechanisms by which SlJAZ11 can induce regeneration.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Characterization of JAZ-interacting bHLH transcription factors that regulate jasmonate responses in Arabidopsis.J Exp Bot. 2011 Mar;62(6):2143-54. doi: 10.1093/jxb/erq408. Epub 2011 Feb 14. J Exp Bot. 2011. PMID: 21321051 Free PMC article.
-
A maize jasmonate Zim-domain protein, ZmJAZ14, associates with the JA, ABA, and GA signaling pathways in transgenic Arabidopsis.PLoS One. 2015 Mar 25;10(3):e0121824. doi: 10.1371/journal.pone.0121824. eCollection 2015. PLoS One. 2015. PMID: 25807368 Free PMC article.
-
Jasmonate regulates leaf senescence and tolerance to cold stress: crosstalk with other phytohormones.J Exp Bot. 2017 Mar 1;68(6):1361-1369. doi: 10.1093/jxb/erx004. J Exp Bot. 2017. PMID: 28201612 Review.
-
JAZ repressors and the orchestration of phytohormone crosstalk.Trends Plant Sci. 2012 Jan;17(1):22-31. doi: 10.1016/j.tplants.2011.10.006. Epub 2011 Nov 21. Trends Plant Sci. 2012. PMID: 22112386 Review.
-
The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana.Plant Cell. 2011 May;23(5):1795-814. doi: 10.1105/tpc.111.083261. Epub 2011 May 6. Plant Cell. 2011. PMID: 21551388 Free PMC article.
Cited by
-
The AlkB Homolog SlALKBH10B Negatively Affects Drought and Salt Tolerance in Solanum lycopersicum.Int J Mol Sci. 2023 Dec 22;25(1):173. doi: 10.3390/ijms25010173. Int J Mol Sci. 2023. PMID: 38203345 Free PMC article.
-
Transcriptomic and metabolomic profiling provide insight into the role of sugars and hormones in leaf senescence of Pinellia ternata.Plant Cell Rep. 2024 Apr 22;43(5):125. doi: 10.1007/s00299-024-03222-x. Plant Cell Rep. 2024. PMID: 38647720
-
Genome-wide annotation and expression analysis of WRKY and bHLH transcriptional factor families reveal their involvement under cadmium stress in tomato (Solanum lycopersicum L.).Front Plant Sci. 2023 Jan 25;14:1100895. doi: 10.3389/fpls.2023.1100895. eCollection 2023. Front Plant Sci. 2023. PMID: 36760632 Free PMC article.
-
Genome-wide insights into the nomenclature, evolution and expression of tobacco TIFY/JAZ genes.Planta. 2025 Apr 4;261(5):103. doi: 10.1007/s00425-025-04676-3. Planta. 2025. PMID: 40183817
-
The CALCINEURIN B-LIKE 4/CBL-INTERACTING PROTEIN 3 module degrades repressor JAZ5 during rose petal senescence.Plant Physiol. 2023 Sep 22;193(2):1605-1620. doi: 10.1093/plphys/kiad365. Plant Physiol. 2023. PMID: 37403193 Free PMC article.
References
-
- Lim P, Kim H, Gil N. Leaf Senescence. Annu Rev Plant Biol. 2007; 58(1): 115–136. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
