

The meiotic cohesin subunit REC8 contributes to multigenic adaptive evolution of autopolyploid meiosis in Arabidopsis arenosa
- PMID: 35830475
- PMCID: PMC9312919
- DOI: 10.1371/journal.pgen.1010304
The meiotic cohesin subunit REC8 contributes to multigenic adaptive evolution of autopolyploid meiosis in Arabidopsis arenosa
Abstract
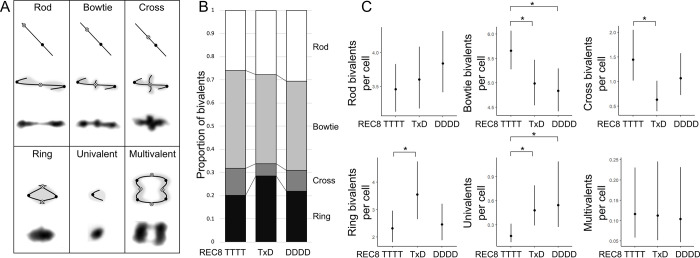
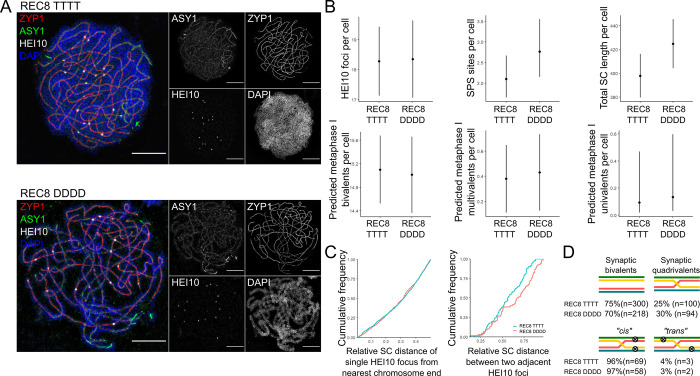
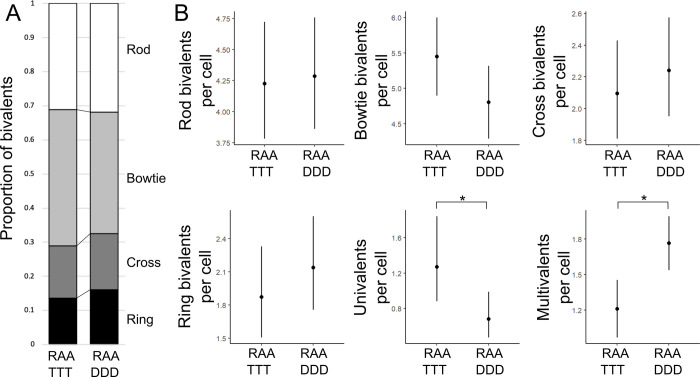
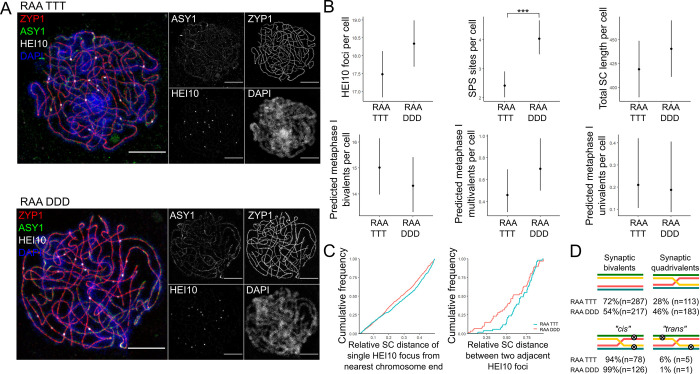
Genome duplication, which leads to polyploidy, poses challenges to the meiotic segregation of the now-multiple homologous chromosome copies. Genome scan data showed previously that adaptation to polyploid meiosis in autotetraploid Arabidopsis arenosa is likely multigenic, involving genes encoding interacting proteins. But what does this really mean? Functional follow-up studies to genome scans for multigenic traits remain rare in most systems, and thus many mysteries remain about the "functional architecture" of polygenic adaptations. Do different genes all contribute subtle and additive progression towards a fitness optimum, or are there more complex interactions? We previously showed that derived alleles of genes encoding two interacting meiotic axis proteins (ASY1 and ASY3) have additive functional consequences for meiotic adaptation. Here we study derived versus ancestral alleles of the meiotic cohesin subunit REC8, which has roles in chromatin condensation, recruiting the axes, and other critical functions in meiosis. We use genetic and cytological approaches to assess the functional effects of REC8 diploid versus tetraploid alleles, as well as their interaction with ancestral versus derived alleles of ASY1 and ASY3. We show that homozygotes for derived (tetraploid) REC8 alleles have significantly fewer unpaired univalents, a common problem in neotetraploids. Interactions with ASY1 and ASY3 are complex, with the genes in some cases affecting distinct traits, and additive or even antagonistic effects on others. These findings suggest that the road to meiotic adaptation in A. arenosa was perhaps neither straight nor smooth.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Derived alleles of two axis proteins affect meiotic traits in autotetraploid Arabidopsis arenosa.Proc Natl Acad Sci U S A. 2020 Apr 21;117(16):8980-8988. doi: 10.1073/pnas.1919459117. Epub 2020 Apr 9. Proc Natl Acad Sci U S A. 2020. PMID: 32273390 Free PMC article.
-
A novel allele of ASY3 is associated with greater meiotic stability in autotetraploid Arabidopsis lyrata.PLoS Genet. 2020 Jul 15;16(7):e1008900. doi: 10.1371/journal.pgen.1008900. eCollection 2020 Jul. PLoS Genet. 2020. PMID: 32667955 Free PMC article.
-
Selection on meiosis genes in diploid and tetraploid Arabidopsis arenosa.Mol Biol Evol. 2015 Apr;32(4):944-55. doi: 10.1093/molbev/msu398. Epub 2014 Dec 26. Mol Biol Evol. 2015. PMID: 25543117 Free PMC article.
-
Many functions of the meiotic cohesin.Chromosome Res. 2010 Dec;18(8):909-24. doi: 10.1007/s10577-010-9169-0. Epub 2010 Nov 18. Chromosome Res. 2010. PMID: 21086039 Review.
-
Meiotic prophase-like pathway for cleavage-independent removal of cohesin for chromosome morphogenesis.Curr Genet. 2019 Aug;65(4):817-827. doi: 10.1007/s00294-019-00959-x. Epub 2019 Mar 28. Curr Genet. 2019. PMID: 30923890 Review.
Cited by
-
The plant early recombinosome: a high security complex to break DNA during meiosis.Plant Reprod. 2024 Dec;37(4):421-440. doi: 10.1007/s00497-024-00509-7. Epub 2024 Sep 27. Plant Reprod. 2024. PMID: 39331138 Free PMC article. Review.
-
The synaptonemal complex stabilizes meiosis in allotetraploid Brassica napus and autotetraploid Arabidopsis thaliana.New Phytol. 2025 Apr;246(2):581-597. doi: 10.1111/nph.70015. Epub 2025 Feb 18. New Phytol. 2025. PMID: 39963072 Free PMC article.
-
Genetic factors inherited from both diploid parents interact to affect genome stability and fertility in resynthesized allotetraploid Brassica napus.G3 (Bethesda). 2023 Aug 9;13(8):jkad136. doi: 10.1093/g3journal/jkad136. G3 (Bethesda). 2023. PMID: 37313757 Free PMC article.
-
The genetic consequences of range expansion and its influence on diploidization in polyploids.bioRxiv [Preprint]. 2024 Apr 17:2023.10.18.562992. doi: 10.1101/2023.10.18.562992. bioRxiv. 2024. Update in: Am Nat. 2025 Feb;205(2):203-223. doi: 10.1086/733334. PMID: 37905020 Free PMC article. Updated. Preprint.
-
Chromosomal Instability in Genome Evolution: From Cancer to Macroevolution.Biology (Basel). 2023 Apr 28;12(5):671. doi: 10.3390/biology12050671. Biology (Basel). 2023. PMID: 37237485 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
